Download

1 / 92
980 likes | 1.05k Views
Explore the maturation process of B and T lymphocytes, including gene rearrangement, receptor expression, selection events, and differentiation into distinct subpopulations. Learn about V(D)J recombination in developing lymphocytes and its role in the adaptive immune system.

E N D
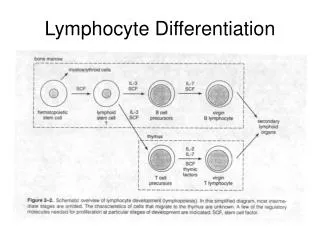
OVERVIEW OF LYMPHOCYTE DEVELOPMENT • The maturation of B and T lymphocytes involves a seriesof events that occur in the generativelymphoid organs .These events include the following: • Commitment of progenitor cells to the B lymphoid orT lymphoid lineage. • Proliferation of progenitors and immature committedcells at specific early stages of development,providing a large pool of cells that can generate usefullymphocytes. • The sequential and ordered rearrangement of antigenreceptor genes and the expression ofantigen receptorproteins. (The terms rearrangement and recombinationare used interchangeably.)
OVERVIEW OF LYMPHOCYTE DEVELOPMENT • Selection events that preserve cells that have producedfunctional antigen receptor proteins and eliminatepotentially dangerous cells that strongly recognizeself antigens. These checkpoints during developmentensure that lymphocytes that express functional receptorswith useful specificities will mature and enter theperipheral immune system. • Differentiation of B and T cells into functionally andphenotypically distinct subpopulations. B cells developinto follicular, marginal zone, and B-1 cells; and T cellsdevelop into CD4+ and CD8+ αβ T lymphocytes, naturalkiller T (NKT) cells, MAIT cells, and γδ T cells.
B-Cell Development in the Primary Lymphoid Organs • Commitment to the B-cell lineage reflects differential activation of transcription factors that progressively lock the cell into the B-cell pathway. • B-cell development is typically viewed as a linear, stepwise process that is focused on the assembly and testing of immunoglobulin function, first in the fetal liver and bone marrow and then in the periphery: • Failure to assemble a functional receptor leads to cell death. • Expression of a functional receptor subjects the B cell to antigen selection. • B cells with inappropriate specificities tend to be eliminated. • B cells responding appropriately to external antigen can develop either into immunoglobulin-secreting plasma cells or into memory cells.
V(D)J recombination V(D)J recombination is the unique mechanism of genetic recombination that occurs only in developing lymphocytes during the early stages of T and B cell maturation. It involves somatic recombination, and results in the highly diverse repertoire of antibodies/immunoglobulins (Igs) and T cell receptors (TCRs) found on B cells and T cells, respectively. The process is a defining feature of the adaptive immune system. V(D)J recombination occurs in the primary lymphoid organs (bone marrow for B cells and thymus for T cells) and in a nearly random fashion rearranges variable (V), joining (J), and in some cases, diversity (D) gene segments. Simplistic overview of V(D)J recombination of immunoglobulin heavy chains
Mechanism: Heavy chain • In the developing B cell, the first recombination event to occur is between one D and one J gene segment of the heavy chain locus. Any DNA between these two gene segments is deleted. • This D-J recombination is followed by the joining of one V gene segment, from a region upstream of the newly formed DJ complex, forming a rearranged VDJ gene segment. All other gene segments between V and D segments are now deleted from the cell’s genome. • Primary transcript (unspliced RNA) is generated containing the VDJ region of the heavy chain and both the constant mu and delta chains (Cμ and Cδ). (i.e. the primary transcript contains the segments: V-D-J-Cμ-Cδ). • The primary RNA is processed to add a polyadenylated (poly-A) tail after the Cμ chain and to remove sequence between the VDJ segment and this constant gene segment. Translation of this mRNA leads to the production of the Ig M heavy chain protein
Mechanism: light chain • The kappa (κ) and lambda (λ) chains of the immunoglobulin light chain loci rearrange in a very similar way, except that the light chains lack a D segment. • In other words, the first step of recombination for the light chains involves the joining of the V and J chains to give a VJ complex before the addition of the constant chain gene during primary transcription. • Translation of the spliced mRNA for either the kappa or lambda chains results in formation of the Ig κ or Ig λ light chain protein. • The immunoglobulin lambda light chain locus contains protein-coding genes that can be lost with its rearrangement. • Assembly of the Ig μ heavy chain and one of the light chains results in the formation of membrane bound form of the immunoglobulin IgM that is expressed on the surface of the immature B cell.
Junctional diversity RAG: the recombination-activating gene encode enzymes that play an important role in the rearrangement and recombination of the genes of immunoglobulin. This gene expression is restricted to lymphocytes during their developmental stages. TdT: the terminal deoxynucleotidyl transferase is a specialized DNA polymerase expressed in immature, pre-B, pre-T lymphoid cells. TdT adds N-nucleotides to the V, D, and J exons of the TCR and BCR genes during antibody gene recombination, enabling the phenomenon of junctional diversity.
B-Cell Development in the Periphery • T cell–independent activation of naïve B cells results in terminal differentiation into short-lived plasma cells. • T cell–dependent activation of B cells: • Induces germinal center formation, permitting somatic hypermutation and class-switch recombination (CSR) • Results in differentiation into high-affinity memory B cells (reactive memory) and plasma cells secreting high-affinity antibodies (protective memory) • Generates long-term humoral immune protection
B-Cell Development in the Periphery • The longevity of plasma cells is supported by highly specialized survival niches in bone marrow. • T-follicular helper (Tfh) cells control late B-cell differentiation by cell bound ligands and secreted cytokines. • Activated B cells control T-cell development by presentation of antigen and co-stimulation.
Early T-Cell Development • • Peripheral T cells are the progeny of hematopoietic stem cells (HSCs) from bone marrow or fetal liver. • • T-cell development occurs in and requires the thymus. • • Patients without a thymus as a result of FOXN1 mutations or DiGeorge syndrome lack circulating T cells. • • Early thymic progenitors (ETPs) are the first cells to seed the thymus.
Early T-Cell Development • • T-cell lineage commitment requires: • • Notch signals • • Upregulation of lineage-defining transcription factors • • Progression through the double negative (DN) stage • • T-cell formation requires production of functional αβ or γδ T-cell receptor (TCR)
The Notch family of proteins are cell surface molecules that are proteolytically cleaved when they interact with specific ligands on neighboring cells. • The cleaved intracellular portions of Notch proteins migrate to the nucleus and modulate the expression of specific target genes. • Notch-1 , a member of the Notch family, is activated in lymphoid progenitor cells and collaborates with a transcription factor called GATA3 to commit developing lymphocytes to the T lineage.
Schematic of counter balancing and dose-dependent regulatory forces that act on T-cell precursors. Regulators named in red denote forces diverting cells from the T-cell pathway. Of these, only EBF and Pax5 are not normally active in the thymus during these stages. Notch signaling acts as a constraint on all of these alternatives. This dynamic equilibrium is normally resolved at the DN3 stage, in part by the downregulation of PU.1, but almost certainly through other regulatory changes as well.
Commitment to The T Cell Lineagesand Proliferation of Progenitors • In humans, IL-7 is required for the proliferation of T cell progenitors but not of progenitors in the B lineage. • IL-7 is produced by stromal cells in the bone marrow and by epithelial and other cells in the thymus.
Commitment to The T Cell Lineagesand Proliferation of Progenitors • Mutations in a chain of the IL-7 receptor, give rise to X-linked severe combined immunodeficiency disease (X-SCID). • This disease is characterized by a block in T cell and NK cell development, but normal B cell development, reflecting the role of IL-7.
MicroRNAs And Lymphocyte Development • Beside the role of transcription factors in lymphocytes gene expression, an additional level of regulation is mediated by microRNAs (miRNAs). • miRNAs are small endogenous noncoding RNAs that are initially generated in the nucleus as longer primary miRNA transcripts that are processed at this site by an endoribonuclease called Drosha into shorter premiRNAs that have a stem loop structure and can be exported into the cytosol. • In the cytosol, the pre-miRNA is processed by another endoribonuclease called Dicer into a short double-stranded miRNA about 21 to 22 base pairs in length, one strand of which can be used to pair with a complementary sequence in a number of cellular mRNAs.
These mRNAs associate with miRNAs and proteins called Argonaute proteins to form complexes known as RISC (RNA-induced silencing complex) . If the 6- to 8-base pair miRNA seed sequence is not perfectly complementary to the mRNA, the mRNAis prevented from being translated efficiently. Otherwise they are targeted for degradation.
T-CELL RECEPTOR REARRANGEMENT AND β SELECTION • One key step in T-cell lineage commitment is rearrangement of a functional TCR, be it an αβ or γδ TCR. • Rearrangement of the TCR gene loci within the developing thymocyte is a highly ordered sequence of recombination events mediated by the recombinase activating gene (RAG) enzymes. • RAG proteins are induced early in DN thymocytes and initiate recombination of the TCR γ, δ, and β genes. VDJ (or VJ) recombination of these loci must yield successful, in-frame rearrangements or the cell will stall at this stage and die.
γδ T cells • For cells of the γδ T-cell lineage, signaling via the newly rearranged γδ TCR complex directs the maturation and export of the γδ T cell into the periphery. • This developmental checkpoint is critical for the selection of the γδ T-cell repertoire and, if unsuccessful, the thymocytes may either die or be redirected to the αβ lineage.
αβ T cells • For cells that will enter the αβ T-cell compartment, the developmental checkpoints are distinct from that of γδ T cells. • Upon productive rearrangement of the TCR β gene, this protein will pair with preT α chain (surrogate α chain) to create the pre-TCR complex. • The pre-TCR complex associates with the CD3 molecules and induces constitutive signaling within the cells.
β Selection and the Appearance of Double-Positive Thymocytes • Signaling by the pre-TCR complex in DN thymocyte facilitates the process of β selection, which instructs the cell to undergo a burst of proliferation and suppresses further β−chain rearrangement. After β selection, the thymocytes will begin to coexpress both CD4 and CD8 on the cell surface. • These cells are termed DP cells. They subsequently initiate recombination of the TCR α gene loci. As a result of the organization of the TCR α loci, DP thymocytes have multiple opportunities to generate a successful TCR α rearrangement. • Consequently most DP thymocytes express an αβ TCR. Only a fraction of these cells will progress into the periphery as CD4 or CD8 SP T cells.
Double-Negative (DN), Double-Positive (DP), and Single-Positive (SP) Thymocytes • Progressive stages of thymocyte development are identified by the expression profiles of CD4 and CD8 coreceptors. • DN (CD4−CD8−) thymocytes give rise to DP (CD4+CD8+) and then SP (CD4+CD8− or CD4−CD8+) thymocytes. • CD4 and CD8 T-cell lineage determination is coordinated by the major histocompatibility complex (MHC) recognition of the T-cell receptor (TCR). • MHC class II restriction leads to CD4 T cells, and MHC class I restriction leads to CD8 thymocytes, which then leave the thymus to become mature CD4 or CD8 T cells.
Positive Selection and Negative Selection • Positive selection promotes the survival of double-positive (DP) thymocytes whose αβ T-cell receptor (TCR) can interact with MHC–self peptide complexes expressed by the thymic cortical epithelial cells. • Negative selection deletes self-reactive T cells from the repertoire by inducing apoptosis in thymocytes expressing an αβ TCR with high affinity for self peptides, promoting central tolerance.
How Does Selection Occur? • Thymic selection involves multiple interactions of DP and SP (single positive) thymocytes with both the cortical and medullary thymic stromal cells, as well as dendritic cells and macrophages. • Selection results in a mature T-cell population that is both self-MHC restricted and self-tolerant. • DP thymocytes that express new TCR dimers browse the MHC/ peptide complexes expressed by the cortical thymic epithelial cells (cTECs). • The large majority of DP thymocytes die in the cortex by neglect because of their failure to bind MHC/peptide combinations with sufficient affinity. • The small percentage whose TCRs bind MHC/ peptide with high affinity die by clonal deletion (negative selection).
How Does Selection Occur? • Those DP thymocytes whose receptors bind to MHC/peptide with intermediate affinity are positively selected and mature to single positive (CD4 or CD8) T lymphocytes. • These migrate to the medulla, where they are exposed to AIRE medullary thymic epithelial cells (mTECs), which express tissue-specific antigens and can mediate negative selection. • Medullary dendritic cells can acquire mTEC antigens by engulfing mTECs, and mediate negative selection (particularly of MHC Class II restricted CD4 thymocytes).
Relationship between TCR affinity and selection. • The previous figure schematically illustrates the association between thymocytes’ fate and the affinity of their TCR for self-MHC/peptide complexes that they encounter in the thymus. • Fewer than 5% of thymocytes produce TCRs that bind to MHC/peptide complexes with high affinity. Most of these will be deleted by negative selection (some will become regulatory T cells and other specialized cell types). • More than 90% generate TCRs that either do not bind to MHC/ peptide complexes or bind them with very low affinity. These die by neglect. • Fewer than 5% generate TCRs that bind with just the right intermediate affinity to self-MHC/peptide complexes. These will survive and mature.
In humoral immune responses, B lymphocytes are activated by antigen and secrete antibodies that act to eliminate the antigen. Both protein and nonprotein antigens can stimulate antibody responses. • B cell responses to protein antigens require the contribution of CD4+ helper T cells specific for the antigen. • Helper T cell-dependent B cell responses to protein antigens require initial independent activation of naive T cells in the T cell zones and of B cells in lymphoid follicles in lymphoid organs, each specific for a different part of the same protein antigen.
A B cell that recognizes a specific epitope on the protein antigen internalizes the protein, processes it, and exhibits a specific peptide epitope on its MHC class II molecules.