Download

1 / 43
430 likes | 688 Views
B-Lymphocyte Activation and Function I Structural Basis of Antibody Diversity Lecture C4-09M. Reading in The Immune System by Parham: Chapter 2 (pages 31-47) Helpful web sites: http://www.classwire.com/Login (website for Text Book)

E N D
B-Lymphocyte Activation and Function IStructural Basis of Antibody DiversityLecture C4-09M • Reading in The Immune System by Parham: Chapter 2 (pages 31-47) • Helpful web sites: • http://www.classwire.com/Login (website for Text Book) • http://www.medicine.dal.ca/micro/education/pimunit/home.htm • http://www.whfreeman.com/kuby/ (has good self test) • http://www-micro.msb.le.ac.uk/MBChB/ImmGloss.html (Glossary of terms)
INNATE VS. ADAPTIVE IMMUNITY • Innate immunity • involves largely phagocytic cells (granulocytes and macrophages) and provides a first line of defense. • Innate immunity deals with the diversity of pathogens that a person might encounter in their lifetime by having available (at all times) a few types of recognition molecules, each of which recognize a large number of pathogens. • Adaptive immunity • is stimulated by exposure to infectious agents and increases in magnitude and defensive capability with each successive exposure to a particular pathogen. • Adaptive immunity is mediated by lymphocytes – • B-lymphocytes (also called B-cells) and T-lymphocytes (also called T-cells).
ADAPTIVE IMMUNITY • B-cells (along with T-cell help) synthesize antibodies that mediate humoral immunity and T-cells are major players in cellular immunity. • The mechanisms of adaptive immunity are ones that improve recognition rather than destruction. • Somatic gene rearrangement and somatic mutation in the genes for antigen receptors provide the lymphocyte population with a set of highly diverse antigen receptors, which collectively have the ability to recognize a vast array of antigens. • An individual lymphocyte expresses receptors of a single and unique antigen-binding specificity.
ADAPTIVE IMMUNITY • B cells and T cells recognize different types of antigens. B-cell receptors bind whole molecules and intact pathogens, whereas T cell receptors only interact with pathogen-derived peptides bound to proteins of the major histocompatibility complex. • IMMUNOLOGICAL MEMORY – Both B cells and T cells of the adaptive immune system “remember” each encounter with a pathogen or foreign antigen, so that subsequent encounters stimulate increasingly more effective defense mechanisms.
Acquired/Adaptive Immunity • Acquired immunity develops during a host’s lifetime. • Six major characteristics of acquired immunity are: • Specificity • Self-nonself discrimination • Inducible • Diversity • Memory • Self-limiting
STRUCTURAL BASIS OF ANTIBODY DIVERSITY • Antibodies are the antigen-specific products of B cells, and the production of antibody in response to infection is the main contribution of B cells to adaptive immunity. • The antibody molecule has two separate functions: • one is to bind specifically to molecules from the pathogen that elicited the immune response; • the other is to interact with various cells and molecules to destroy the pathogen once the antibody is bound to it. • These functions are structurally separated in the antibody molecule, one part of which specifically recognizes antigen and the other engages effector mechanisms that dispose of it. • The antigen-binding region varies extensively between antibody molecules and thus is known as the variable region or V region. • The region of the antibody molecule that engages the effector functions of the immune system does not vary in the same way and is thus called the constant region or C region.
THE IMMUNOGLOBULIN G (IgG) MOLECULE Note: 2 identical heavy chains (green) and 2 identical light chains (yellow). Hinge Note: Location of the variable (red) and constant (blue) regions in the antibody molecule. Concept: Antibodies are composed of polypeptides with variable and constant regions.
Concept: The antibody molecule can readily be cleaved in to functionally distinct fragments. Note: Papain cleaves the IgG into three pieces: two Fab (Fragment antigen binding) fragments and one Fc fragment (Fragment crystalizable). Note: Pepsin cleaves IgG into one (Fab’)2 fragment. The Fc is broken into several smaller fragments.
Concept: All classes of antibody have the same basic structure. Two identical heavy chains and two identical light chains: each of which contributes to antigen binding. The Structural Organization of the Human Immunoglobulin Isotypes.
HEAVY CHAINS • All heavy chains are made up of discrete domains. • The amino terminal domain, of VH, displays the greatest variability. • The remaining domains, of CH, of the heavy chain form the constant region. • There are five classes of heavy chains designated: γ, α, µ, δ, ε. • γ and α are further divided into subclasses – γ1, γ2, γ3, γ4; and α1 & α2 – thus …. • There are nine isotypes of antibodies as defined by the heavy chains. • All heavy chains are expressed in one of two molecular forms, fluid phase and membrane bound forms.
Light CHAINS • All antibody light chains fall into two classes – • κ (kappa) and λ (lambda). • Each light chain, whether κ or λ, is folded into separate V and C domains. • There is a single gene for the kappa constant region, thus there is a single isotype of kappa chains. • There are six separate genes for the constant region of lambda chains, thus, there are six isotypes of lambda chain.
Immunoglobulin Domains • Both H and L chains contain several homologous units of about 110 amino acids termed a – DOMAIN. • Each domain contains an intrachain disulfide bond which forms a loop of approximately 60 amino acids. • L chains contain one variable domain (VL), and one constant domain (CL) • H chains contain one variable domain – (VH) – and either three or four constant domains – (CH1, CH2, CH3, and CH4) depending on the antibody class.
General Structures of the Five Classes of Immunoglobulins in Serum IgE IgG IgD IgA (dimer) IgM (pentamer) Note: The heavy and light chains of an immunoglobulin molecule are made up of similar protein domains.
Immunoglobulin Isotypes are Selectively Distributed in the Body • The principal immunoglobulins in plasma are IgG, IgA and IgM (the concentration is IgG>IgA>IgM. In a healthy host IgM is found only in the plasma, whereas IgG and IgA are found in other compartments. • IgG and monomeric IgA are the major isotypes in the extracellular fluid. • IgG is the only isotype to cross the placenta. • Dimeric IgA is the predominate isotype in secretions across epithelia. • IgE is associated mainly with mast cells and basophiles (subject of a later lecture). • IgD is predominately found as a receptor on B cells. • In healthy individuals the Ig concentration in the CNS is low.
Concept: An antigen-binding site is formed from the hypervariable regions of a H-chain and L-chain V domain. • The amino-terminal domain of both H and L chains is termed a variable (V) region due to the discovery of extensive seequence divergence between different antibody proteins in this part of the molecule. These are designated VH and VL for H and L chains, respectively. • V regions have been demonstrated to be responsible for antigen specificity of the immunoglobulin. • Sequence analysis of many VH and VLdomains revealed that variability is concentrated in several hypervariable (HV) regions. • There are three hypervariable regions in each VH and VL domain. • The stretches of amino acid between the HVs are termed framework regions and these exhibit far less variability. • The interaction between the HVs of the H and L chains form the antigen binding site of the antibody molecule. • Since these HVs fold into a structure that is complementary to the antigen epitope, the HVs are also called complementarity-determining regions (CDRs).
Antigen Combining Site A L-chain HVs • Schematic of and IgG molecule showing the approximate location of the hypervariable regions (CDRs). • Schematic showing how the CDRs in each heavy- and light-chain pair for an antigen binding site. H-chain HVs • 1. Hypervariable Regions (HV): areas of extreme variability in the V domain of both H and L chains. • Hypervariable regions are also called complementarity determining regions (CDRs). • Framework Regions: stretches of amino acids between the hypervariable regions that show relatively less variability. B
Antigen Receptor Diversity • The human genome is estimated to contain 34 thousand genes (give or take a few thousand) • The estimated number of antibody specificities is estimated to be 30 million or more; the number of T cell receptor specificities is about the same. • Is there a problem with these numbers????? • How can fewer than 105 genes encode for nearly 108 antigen receptor specificities plus all of the other proteins needed by the body???????? • The answer – • Each receptor chain is encoded by different gene segments
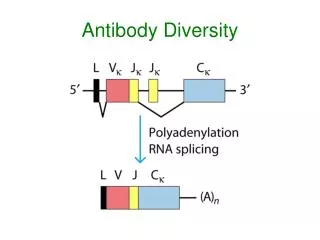
CONCEPT: The DNA sequence encoding a complete V region is generated by the somatic recombination of separate gene segments. • Three separate multigene families (kappa, lambda, and heavy chain loci) located on different chromosomes encode immunoglobulin light chains and heavy chains. • In germline DNA, each multigene family contains numerous gene segments • Ig genes undergo a process of somatic DNA recombination or rearrangement during B cell development to form the variable regions of light chains and heavy chains. • Functional light-chain and heavy-chain genes are generated by random rearrangement of the variable region gene segments (two for light chains and three for heavy chains) in developing B lineage cells.
CONCEPT: Light chain variable regions are assembled from to gene segments – V and J (for joining).
CONCEPT: Heavy chain variable regions are assembled from three gene segments – V (variable), D (for diversity) and J (for joining).
CONCEPT: The reaction that recombines V,D, and J gene segments involves both lymphocyte-specific and ubiquitous DNA-modifying enzymes that are collectively call the recombinase complex. • Rearrangement of Ig genes occurs principally by a mechanism of excision (looping out) of the DNA between the various gene segments followed by ligation of these segments. • Ig gene rearrangement is a special kind of recombination involving non-homologous gene segments that is mediated by a system of enzymes. • The enzymes involved include: • lymphocyte-specific enzymes, which are products of the recombinase activating genes (Rag1 and Rag2) • and, other nuclear DNA-modifying enzymes (DNA repair) that are not tissue specific.
RECOMBINASES • Recombinases (RAG-1 and RAG-2) recognize specific DNA sequences called recombination signal sequences (RSS) that flank each V, D, and J gene segment. • Recombinases are cell type-specific. They are active in lymphocytes but not in non-lymphoid cells. • Recombinases function in the early developmental stages of B and T lymphocytes and are absolutely required for recombination to occur. • Recombinase activating genes 1 and 2 (Rag-1 and Rag-2) are present in all cells but are only expressed in B and T cells.
CONCEPT: Gene segments encoding the variable region are joined by recombination at recombination signal sequences (RSS).
CONCEPT: The diversity of the immunoglobulin repertoire is generated by four main processes. • Multiple inherited germline gene segments used in different combinations. • Junctional diversity — antibody molecules show the greatest variability at the junctions of the variable gene segments which form the third hypervariable region, or CDR3. Junctional diversity arises by two mechanisms: • Introduction of P-nucleotides by a feature of the recombination reaction that generates short palidromic sequences at the cut ends of the DNA strands. • N-nucleotides formed from the untemplated addition of nucleotides by the enzyme terminal deoxynucleotidyl transferase (TdT). • Combination of any H-chain with any L-chain. • Somatic Hypermutation (occurs after encounter with antigen).
CONCEPT: Junctional diversity is generated during recombination of variable gene segments.
CONCEPT: The development of B cells proceeds through stages defined by the rearrangement and expression of the immunoglobulin genes.
The Other Side of the Coin • Antigens/Immunogens • Antigen – antibody generators • Immunogen – any substance that can elicit an immune response in either B cells or T cells
ANTIGENS/Immunogens • Substances capable of inducing an immune response are called antigens or immunogens. • To protect against infectious disease, the immune system must be able to recognize bacteria, bacterial products (e.g., toxins), fungi, parasites, and viruses as immunogens. • The immune system actually recognizes particular macromolecules of an infectious agent, generally either proteins or polysaccharides. • Proteins are the most potent immunogens with polysaccharides ranking second. • In contrast, lipids and nucleic acids of an infectious agent generally do not serve as immunogens unless they are complexed to proteins or polysaccharides.
Contribution of the Immunogen to Immunogenicity • FOREIGNNESS – • In order to elicit an immune response, a molecule must be recognized as nonself by the biological system (i.e., the host). • MOLECULAR SIZE – • There is a correlation between the size of a macromolecule and its immunogenicity. Generally, a molecule must have a molecular mass >10,000 daltons. • CHEMICAL COMPOSITION AND COMPLEXITY – • Size and foreignness are not, by themselves, sufficient to make a molecule immunogenic – other properties are needed. The macromolecules must have internal complexity. Simple polymers are not immunogenic. • SUSCEPTIBILITY TO ANTIGEN PROCESSING AND PRESENTATION – • The development of most humoral and all cell-mediated immune responses requires interaction of T cells with antigen that has been processed and presented together with MHC molecules. Thus, if a macromolecule can not be degraded by the biological system it is not immunogenic.
Contribution of the Biological System to Immunogenicity • Even if a macromolecule has the properties that contribute to immunogenicity, its ability to produce an immune response will depend on certain properties of the biological system that the antigen encounters. • GENOTYPE OF THE RECIPIENT HOST – • The genetic composition of the immunized host influences the type and degree of the manifested response. • HEALTH AND WELL BEING OF THE HOST – • sickness, stress, malnutrition, and age (too old or too young) of the host can affect the type and degree of an immune response. • IMMUNOGEN DOSE AND ROUTE OF ADMINISTRATION – • Too high or too low of a dose can result in a poor immune response (tolerance to the antigen). The administration route strongly influences which immune organs and cell populations will be involved in the response.
EPITOPES • The molecular sites that stimulate immune responses are termed epitopes or antigenic determinants • B-cell Epitopes — may be sequential or conformational • B cell epitopes can be found in proteins, carbohydrates, lipids, chemicals, drugs, etc. • Haptens --Non-immunogenic substances (small drugs, lipids,peptides, etc) can be coupled to carrier proteins and then can serve as B cell epitopes. • B cells recognize antigen that is free in the fluid phase. • T-cell Epitopes — are made up of sequential residues in a protein immunogen. • T cells only recognize antigen after it has been processed by APCs and presented in the context of MHC
CONCEPT: Epitopes of a protein can be linear or discontinuous (conformational) A linear epitope of a protein antigen is formed from contiguous amino acids. A discontinuous epitope is formed from amino acids from different parts of the polypeptide that are brought together when the chain folds, i.e., are dependent on the conformation of the protein.
Concept: Epitopes can bind in pockets or grooves or on extended surfaces in the binding site of antibodies.
Forces Binding Antigen to Antibody • The combination of antigen with antibody is a non-covalent interaction. Ag + Ab Ag-Ab (immune complex) • The strength of the association is a summation of several weak intermolecular forces: • Van der Waal’s Forces • Hydrogen Bonds • Hydrophobic interactions • Electrostatic (Ionic) interactions
Cross-Reactivity of Antibodies • ENIGMA – If antibodies are so highly specific why is it often observed that they will cross-react with unrelated antigens? • Same molecular structure (structurally similar epitope) • Structural mimicry • Examples • ABO blood group antigens • Rheumatic heart disease • Post-streptoccocal glomerulonephritis
Summary of Antibodies • are glycoproteins • are built of subunits containing • two identical light chains (L chains), each containing about 200 amino acids • two identical heavy chains (H chains), which are at least twice as long as L chains • The first 100 or so amino acids at the N-terminus of both H and L chains vary greatly from antibody to antibody – the are termed the variable (V) regions • unless members of the same clone (often not even then!), no two B cells are likely to secrete antibodies with the same V region • the amino acid sequence variability in the V regions is especially pronounced in 3 hypervariable regions also called CDR’s • the tertiary structure of the antibodies brings the 3 CDR’s of both the H and L chains together to form the antigen binding site which binds the antigen epitope • Only a few different amino acids sequences are found in the C-terminus of the H and L chains and these are called constant (C) regions • two different kinds of C regions for L chains -- kappa (κ) and lambda (λ) • five different kinds of C regions for H chains • mu (µ) chains, IgM • gamma (γ) chains, IgG • alpha (α) chains, IgA • epsilon (ε) chains, IgE • delta (δ) chains, IgD • each of these 5 kinds of H chains can pair with either lambda or kappa L chains
Summary of Antigen Receptor Diversity • Each receptor chain is encoded by several different gene segments • the gene segments are V, D, and J for heavy chains; and V and J for light chains • Each gene segment has an adjacent recombination signal sequenc (RSS) • RSS’s are recognized by two recombinase proteins encoded by Rag-1 and Rag-2 genes • During the differentiation of the B cell the DNA is cut by the recombinase enzymes and recombined to make an intact gene coding for a receptor chain. • DJ joining occurs first in a developing B cells followed by VDJ • VJ recombination of light chain genes occurs after the successful completion of VDJ recombination of the heavy chain genes. • Random assortment of gene segments contributes to diversity • Junctional diversity, P-nucleotides and N-nucleotides contribute to additional receptor diversity. • Somatic hypermutation after the B cell encounters antigen adds additional diversity • Ergo – little bit of DNA – a whole bunch of antibodies