Vertebrate Retinal Histogenesis
600 likes | 798 Views
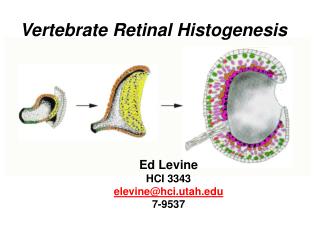
Vertebrate Retinal Histogenesis. Ed Levine HCI 3343 elevine@hci.utah.edu 7-9537. The neural retina contains seven major cell classes derived from the neuroepithelium. Rods Cones. Outer Nuclear Layer (ONL). Horizontal Cells. Muller Glia. Inner Nuclear Layer (INL). Bipolar Cells.

Vertebrate Retinal Histogenesis
E N D
Presentation Transcript
Vertebrate Retinal Histogenesis Ed Levine HCI 3343 elevine@hci.utah.edu 7-9537
The neural retina contains seven major cell classes derived from the neuroepithelium Rods Cones Outer Nuclear Layer (ONL) Horizontal Cells Muller Glia Inner Nuclear Layer (INL) Bipolar Cells Amacrine Cells Ganglion Cells Ganglion Cell Layer (GCL)
Overview of Histogenesis Temporal progression Spatial progression Adult stem cells Regeneration
Overview of Histogenesis Temporal progression Spatial progression Adult stem cells Regeneration
Tissue formation (cell production, histogenesis) from the perspective of developmental time prototypical development of CNS tissue 1 2 3 4 5 6 7 8 developmental time Progenitor cell Post-mitotic differentiated cells
Pulse-chase labeling paradigm Birthdating of differentiated cells Single exposure of 3H-thy (or BrdU) • Fix cells at selected timepoints, stain for BrdU
Birthdating of differentiated cells 3H-thy / BrdU Heavily labeled differentiated cells are considered to be born soon after time of nucleotide labeling
Birthdating of differentiated cells 3H-thy / BrdU • Cells that exit cell cycle are heavily labeled • Cells that continue to proliferate dilute the label and are lightly labeled
2 phases of histogenesis in the rat retina • RGCs, Cones, Horizontal, and Amacrine cells are generated in phase 1 • Amacrine, Rods, Bipolar cells and Muller glia are generated in phase 2 • Many more cells are generated in phase 2
The order of temporal progression is conserved across vertebrates with some exceptions, especially when comparing horizontal cells and cones. • The extent of overlap can vary greatly. In fast developing organisms (frogs and fish), overlap of cell class intervals is extensive. In birds and mammals, overlap is less and is loosely correlated with gestation period. • The relative ratios of cell classes varies between species, especially with respect to rods and cones • In amphibians and fish, histogenesis begins during embryogenesis, but majority of growth occurs posthatch from ciliary margin zone • In birds and mammals, histogenesis is restricted to embryonic and/or early postnatal periods
Retinal histogenesis is more like the second model 1 2 3 4 5 6 7 8 developmental time Non-overlapping cell class generation Overlapping cell class generation
Tissue formation (cell production, histogenesis) from the perspective of developmental time and CC 1 2 3 4 5 6 7 8 developmental time 1 2 3 4 5 6 7 8 9 10 cell cycle #
N-cam Blocking proliferation at early stage of development prior to cell fate commitment does not prevent formation of late cell types in Xenopus embryos treated untreated Pharmacological approach: Hydroxyurea: blocks ribonucleoside diphosphate reductase which converts ribonucleotides to deoxynucleotides, which are necessary for DNA biosynthesis Aphidicolin: inhibitor of DNA Polymerase-alpha photoreceptors amacrines RGCs general neuronal marker Harris and Hartenstein (1991) Neuron 6:499-515
Cell fate is not dependent on the number of CCs a progenitor cell passes through in Xenopus Normal development 1 2 3 4 5 6 7 8 developmental time 1 2 3 4 5 6 7 8 9 10 cell cycle # Development during blocked cell division 1 2 3 4 5 6 7 8 developmental time 1 cell cycle #
Loss of Chx10 causes severe cell number deficits in the developing retina
The cell number defect of the Chx10 null retina is correlated with changes in retinal progenitor cell (RPC) proliferation and cell cycle dynamics, not differentiation (or cell death) Green et al (2003) Development 130:539-552
The cell number defect of the Chx10 null retina is correlated with changes in retinal progenitor cell (RPC) proliferation and cell cycle dynamics, not differentiation (or cell death)
Model: Chx10 regulates CC time, not developmental time Development in absence of Chx10 1 2 3 4 5 6 7 8 developmental time 1 2 3 4 5 cell cycle # Normal development 1 2 3 4 5 6 7 8 developmental time 1 2 3 4 5 6 7 8 9 10 cell cycle #
Counting cell cycles does not appear to regulate the timing of temporal progression
Are RPCs multipotent or lineage restricted? lineage restricted multipotent
Birthdating of differentiated cells 3H-thy or BrdU Lineage analysis of progenitor cells lineage-restricted progenitor ? retrovirus dextran dye cell-specific Cre with floxed reporter multipotent progenitor
Determination of the lineage of a neuroblast in the rat retina (A) A virus containing a functional -galactosidase gene is injected and after 4-6 weeks the retina is stained for the presence of the enzyme. (B) Stained cells forming a strip across the neural retina, including five rods, a rod terminal (t), and a Muller glia cell (mg). Turner and Cepko (1987) Nature 328: 131
Individual progenitors can give rise to multiple cell classes mouse xenopus Rat: Turner and Cepko (1987) Nature 328: 131 Mouse: Xenopus:
Are postnatal rat RPCs multipotent or lineage restricted? lineage-restricted progenitor ? retrovirus multipotent progenitor
Result: Both lineages were observed lineage-restricted progenitor retrovirus multipotent progenitor
As were 2 cell clones, each cell with a different fate lineage-restricted progenitor AND retrovirus multipotent progenitors
Pax6 is required for the multipotent state of mouse RPCs Conditional ablation of Pax6 in the peripheral retina at the optic cup stage causes severe defects in the generation of major retinal cell types Marquardt et al. (2001) Cell 105: 43
Model of requirement for Pax6 in regulating multipotential state Marquardt et al. (2001) Cell 105: 43
Early cell fate in the vertebrate retina is flexible and influenced by extrinsic factors E15 retina cells labeled with thymidine and mixed with other E15 cells (isochronic) are not as likely to differentiate as rods. Instead, they primarily make retinal ganglion cells (RGC). If the same cells are mixed with P1 cells (heterochronic), they tend to differentiate as rods. Watanabe and Raff (1990) Neuron 4:461 • Reh shows similar results using heterochronic co-cultures of dissociated mouse and rat cells J.Neurobiology (1992) 23:1067-1083 • In contrast, Morrow et al observe no influence of P0 cells on E16 cells with respect to rod differentiation J. Neurosci (1998) 18:3738-3748
Other data supporting the model that cell fate determination is dependent on extracellular environment Watanabe and Raff (1990) Neuron 4:461 Early retinal environment is inhibitory to rod differentiation (fewer late retinal cells expressed opsin if cultured in excess of early retinal cells (not specific to retinal cells - observed with other cells as well) (we don’t know what the late cells become e.g. other late cell type, blocked from maturation, early cell type Belliveau et al (2000) J Neurosci 20:2247-2254 Confirms that early retinal environment is inhibitory to rod differentiation, and also observed an increase in bipolar cells Belliveau and Cepko (1998) Development 124:1119-1131 E16 rat RPCs cultured in excess of P0 cells inhibited amacrine cell production (NEGATIVE FEEDBACK MODEL) James et al (2003) J.Neurosci 23:8193-8203 Used RGC markers to show that late rat RPCs can differentiate into RGCs if exposed to early cellular environment (E3 chick retinal cells/CM) (EXAMPLE OF RPCs HAVING POTENTIAL TO GO BACK IN LINEAGE) Also found that early environment (E3 chick retinal cells/CM) inhibits opsin expression in late RPCs Adler and Hatlee (1989) Science 243:391-393 Repka and Adler (1992) Dev Biol 153:242-2549 Cell type produced by dissociated mitotic cell is not specified intrinsically but rather dictated by environment - isolate chick retinal cells give rise to photoreceptors regardless of age (tested up to E8) - argues for model that photoreceptor is default phenotype
rhodopsin antibody: Control +shh Sonic hedgehog promotes rod differentiation and progenitor proliferation in retinal cells Levine et al., 1997, J.Neuroscience 17: 6277
Other extrinsic factors that regulate cell fate determination in the vertebrate retina: retinoic acid notch/delta FGF activin TGF, laminin neurotrophins (NGF, BDNF, NT-3) CNTF
Differentiated cells feedback to limit cell fate/differentiation mitotic progenitors differentiating neurons & glia
Data supporting the model that cell fate determination is programmed cell intrinsically Reh and Kljavin (1989) J. Neurosci 9:4179-4189 Cell type produced by dissociated mitotic cell is reflective of age that cell is initially isolated - Argues for intrinsic information (used E14 and P1 rat cells) Cayouette et al (2003) Neuron 40:897-904 Clonal analysis of proliferation and differentiation in E16 rat explants and dissociated cells Results: The 2 culture conditions generate very similar clones (in size and composition) even though the environments are very different Alexaides and Cepko (1997) Development 124:1119-1131 Early VC1.1+ rat RPCs generate amacrine and horizontal cells and VC1.1- RPCs generate cones Late VC1.1+ rat RPCs generate amacrine and rod cells These results argue that intrinsic differences correlate with bias of cell output
Model of temporal progression Timing appears to be regulated by changes in both cell-extrinsic and cell-intrinsic factors and responses and by negative feedback mechanisms
Overview of Histogenesis Temporal progression Spatial progression Adult stem cells Regeneration
Histogenesis occurs as a wave of differentiation from the central to peripheral retina
Histogenesis occurs as a wave of differentiation from the central to peripheral retina mitotic progenitors differentiating neurons & glia
Histogenesis occurs as a wave of differentiation from the central to peripheral retina mitotic progenitors differentiating neurons & glia
Major Questions How is neurogenesis initiated? How is wave propogated? Is each wave of differentiation coordinated?
Sonic hedgehog drives a wave of neurogenesis across the retina (similar to the Drosophila retina) Neumann and Nuesslein-Volhard, Science 289: 2137
Overview of Histogenesis Temporal progression Spatial progression Adult stem cells Regeneration
The CMZ recapitulates the cellular and molecular mechanisms of embryonic development Perron et al., 1998
The juvenile chick retina contains a CMZ Reh and Fischer, 2001
mammalian pigmented Ciliary Epithelium: a source of in-vitro derived retinal stem cells Tropepe et al., 2000
Adult fish retina: 3 intrinsic neurogenic cell populations