Download
1 / 34
340 likes | 998 Views
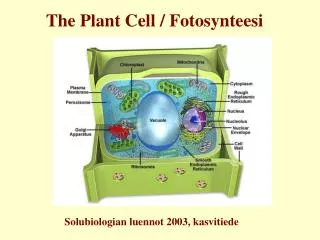
The Plant Cell / Fotosynteesi. Solubiologian luennot 2003, kasvitiede. Fotosynteesissä tuotetaan auringonvalon avulla orgaanisia yhdisteitä. Fotosynteesissä valoenergia muuttuu kemialliseksi energiaksi ja sen avulla CO 2 pelkistyy ja yhtyy H 2 O:stä peräisin olevan vedyn kanssa glukoosiksi.

E N D
The Plant Cell / Fotosynteesi Solubiologian luennot 2003, kasvitiede
Fotosynteesissä tuotetaan auringonvalon avulla orgaanisia yhdisteitä • Fotosynteesissä valoenergia muuttuu kemialliseksi energiaksi ja sen avulla CO2 pelkistyy ja yhtyy H2O:stä peräisin olevan vedyn kanssa glukoosiksi. • 6 H2O + 6 CO2 C6H12O6 + 6O2 • Tämä tapahtuu pitkässä reaktio-ketjussa, jossa energia liitetään vähitellen orgaanisen aineeseen solun rakenteita vahingoittamatta. • Yhteyttämisen vaiheet jaetaan valoreaktioihin (> O2 , NADPH, ATP) & hiilen pelkistykseen, Calvinin sykli (pimeäraktioihin). • Valoreaktiot tylakoidikalvoilla
Valoreaktiot • Klorofyllimolekyylit ja muut pigmenttiaineet etenkin karotenoidit muodostavat valohaavin (200-300 pigmenttimolekyyliä), jonka molekyylit pyydystävät säteilyn energiaa. • Valokvantin eli fotonin osuessa pigmenttimolekyyliin yksi sen elektroneista virittyy eli muuttuu korkeaenergiseksi = siirtyy enemmän energiaa sisältävälle orbitaalille eli kauemmaksi atomiytimestä • Korkeaenergisestä elektronista energia siirtyy helposti naapurimolekyyliin, jonka elektroni vuorostaan virittyy jne. • Lopulta energia välittyy reaktiokeskuksena toimivalle klorofyllimolekyylille, josta se siirtyy elektroninsiirtäjäentsyymille.
Jos väärä aallonpituus niin valokvantti kulkee pigmentin läpi ilman että sitä absorboidaan. Miksi klorofylli on vihreää ? Jos allonpituus on oikea, oikea määrä energiaa niin valokvantti absorboidaan ja elektroni virittyy (siirtyy uudelle orbitaalille)
Virittynyt tila on epästabiili ja se voi stabiloitua eli elektroni palautuu perustasolleen yksinkertaisimmillaan lämpönä tai emittoitumalla (fluoresenssin avulla) Elektroni voi myös stabiloitua siten, että energia siirtyy kokonaan toiselle läheiselle molekyylille. Tällä menetelmällä (energy transfer) absorboitu valoenergia siirtyy pigmenttimolekyyliltä toiselle. Lopuksi virittynyt molekyyli luovuttaa elektronin elektronin vastaanottaja- molekyylille.
Pääasiallisena antennapigmentteinä kasveilla toimivat klorofyllit. Chl a on sekä antennoissa että reaktiokeskuksissa. Chl b pelkästään antennoissa. Myös karotenoideja on antennoissa. Tyypillisesti kasvien antenna-komplekseissa karotenoidi / totaali klorofylli suhde = 0.5 Kaikki klorofyllimolekyylit tylakoideissa ovat liittyneinä spesifisiin proteiineihin. Kaikki nämä proteiinit koodataan tumassa ja kuljetetaan kloroplastiin. Kun chl liittyy proteiiniin sen absorbtiomaksimi siirtyy kohti punaista spektrin aluetta (alhaisempi energia).
Light-harvesting complex II (LHC II) Pääasiallinen pigmenttejä sitova proteiini kloroplastin kalvoissa on LHC-II = light-harvesting complex II (eli fotosynteesissä valoa keräävä pigmentti-proteiini). On arvioitu, että tämä proteiini muodostaa määrällisesti noin puolet kaikista tylakoidien proteiineista. thylakoid membrane
Lateraalinen heterogeenisuus non-appressed region of grana appressed grana region non-appressed region
Lateraalinen heterogeenisuus Komponentit Tylakoidit % Graana Strooma PSII 85 15 PSI 10 90 Cyt b6f kompleksi 50 50 LHC-II 90 10 ATP syntetaasi 0 100 plastosyaniini 40 60
Kloroplastit kykenevät vaikuttamaan LHC-II:n liittymistä PSII:een ja siten säätelemään valokvanttien jakautumista PSI ja PSII kesken. Kun PSII saa enemmän valoa kuin PSI plastokinoni (liikkuva elektronikantaja PSI ja PSII välillä) muuttuu pääosin pelkistyneeksi, jolloin LHC-II kinaasi aktivoituu ja fosforyloi LHC-II, jonka seurauksena LHC-II liikkuu pois grana-alueelta > valon absorbtio PSII:een vähenee. PSI:n aktivaation seuraksena plastokinoni muuttuu hapettuneemmaksi, LHC-II kinaasi muuttuu vähemmän aktiiviseksi ja fosfataasi defosforyloi LHC-II:n, joka liikkuu grana-alueelle > valon absorbtio PSII:een lisääntyy. LHC-II:n aktiivisuuttasäädellään ns. feedback- systeemillä
PSI ja PSII • PSI on korkeamman aallonpituuden reaktiokeskus P700 on tehokkaampi pitkässä punaisessa valossa ja • PSII toimii tehokkaamin punaisessa valossa johtuen alhaisemman aallonpituuden reaktiokeskuksesta P680. Jotta fotosynteesi olisi tehokasta näiden reaktiokeskusten täytyy toimia yhdessä. • Kloroplasteissa toimii sekä ei-syklinen että syklinen elektroninsiirtoketju.
Kloroplasteissa on sekä syklinen että ei-syklinen elektroninsiirtoketju syklinen ei-syklinen
Ei-syklinen elektronin siirto tuottaa O2, ATP ja NADPH Valossa PSII tuottaa vahvan hapettimen P680+ ja suhteellisen pysyvän pelkistimen plastosemikinonin (QA- ). P680+ ottaa elektronit vedeltä ja plastosemikinoni taas pelkistysvoimansa ansiosta siirtää elektronit useiden elektronikantajien kautta P700:lle. Vapautuneen energian avulla muodostetaan protonigradientti, jota käytetään ATP:n synteesiin. Typen assimilaatio Tioredoksiinin pelkistys +H+> NADPH vahva pelkistin pelkistin hv> vahva hapetin hv>hapetin PC = plastocyanin
Fotosysteemi II:n (PSII) reaktiokeskus PSII sisältää elketroninsiirtoon liittyvien komponenettien lisäksi yli 20 proteiinia (D1 ja D2 sitovat elektronin siitoon liittyviä prosteettisia ryhmiä, CP43 ja CP47 sitovat chl a antenna pigmenttejä ja jotkut liittyvät veden hapetukseen).
PSII:n reaktiokeskuksen D1-proteiini aminopää karboksyylipää
Sykliseen elektroninsiirtoketjuun liittyy vain PSI ja se tuottaa ATP:tä. Syklinen reitti tarvitsee kofaktorin, joka mitä todennäköisimmin on ferredoksiini. Valossa PSI pelkistää ferredoksiinin, joka (sen sijaan että siirtäisi elektronin NADP:lle) reagoi ferredoksiini-plastokinoni oksidoreduktaasin kanssa, jolloin elektronit siirtyvät kinoneille. Cyt b6f hapettaa plastokinonin ja protonin kuljetus kalvon läpi mahdollistuu.
Valosta riippuvaa ATP-synteesiä kloroplasteissa kutsutaan fotofosforylaatioksi (muistuttaa mitokondrioiden oksidatiivista fosforylaatiota) ATPsyntetaasi CF0 toimii protonikanavana, koostuu alayksiköistä I-IV CF1-kompleksi sisältää ATP fosfataasiaktiivisuuden
Ilmaraot (stoma, stomata) ovat kasvin epidermisolukossa olevia pieniä aukkoja, joita reunustavat huulisolut. Niitä on yleensä enemmän lehtien ala- kuin yläpinnalla. Ilmarakojen avautuminen ja sulkeutuminen riippuvat huulisolujen nestejännityksestä eli turgorista. Ilmaraot säätelevät kasvin kaasujen vaihtoa ja veden haihdutuksen määrää, joten niillä on tärkeä merkitys kasvien elintoiminnoille. huulisolut ilmarako epidermisolut Huulisoluihin liittyvien apusolujen sijainnin ja lukumäärän mukaan erotetaan erilaisia ilmarakotyyppejä / laitteita
ilmarako päältä ilmarako lehden poikkileikkauksessa ulkokynsi ulkosarana ulkoeteinen viherhiukkanen keskirako tuma sisäeteinen sisäkynsi sisäsarana ilmalokero huulisolu
Calvinin kierto / C3 kasvit Useimmat kasvit tuottavat 3-hiiliatomisen yhdisteen 3-fosfoglyseraatin (3-PGA) ensimmäisenä pysyvänä tuotteena monivaiheisessa reaktiosarjassa, jossa CO2 muutetaan hiili-hydraateiksi. CO2 reagoi ensin 5-hiilisen yhdisteen ribuloosi-1,5-bisfosfaatin (RuBP) kanssa, jota reaktiota katalysoi ribuloosi bisfosfaatti karboksylaasi / oksygenaasi(Rubisco). Syntyy 6-hiili-atominen välituote, joka heti pilkotaan kahdeksi PGA-molekyyliksi.
Calvinin sykli • Rubisco on ensimmäinen hiilen sidonnasta vastaava entsyymi ja se on yleisin liukoinen proteiini kloroplastissa (ja ehkä biosfäärissä). • Rubisco koostuu 8 suuresta L- (large, 56 kDa) ja 8 pienestä S- (small, 14 kDa) alayksiköstä. L-alayksiköt koodataan kloroplastissa ja S-alayksiköt tumassa. Tumassa koodattavat ovat siis tuotava kloroplastiin, jossa ne kloroplastin stroomassa yhtyvät suuriin alayksikköihin näin muodostaen holoentsyymin. • Calvinin sykli voidaan jakaa kolmeen vaiheeseen 1) karboksylaatioon, 2) pelkistykseen ja 3) regeneraatioon. • Kaksivaiheinen pelkistysvaihe muuttaa 3 PGA:n trioosifosfaatiksi eli glyseraldehydi-3-fosfaatiksi (GAP).
Calvinin syklin pelkistymisvaihe, jossa 3-PGA:n karboksyyliryhmä pelkistyy aldehydiksi Glyseraldehydi-3- fosfaatti (GPA)
Calvinin sykli • Syklin regeneraatiovaiheessa palautetaan RuBP (5-hiilinen sokeri) = useita reaktioita (10 entsyymiä, kts. kaavakuva). • Syklin 13 entsyymiä sijaitsevat stroomassa. Rubiscon lisäksi syklissä on kaksi muuta entsyymiä (sedoheptuloosi-1,7-bisfosfataasi ja fosforibulokinaasi). • Monet syklin entsyymeistä ovat yhteisiä glykolyyttisen reitin kanssa. Koska sekä hiilihydraattien synteesiin ja hajotukseen tarvittavia entsyymejä on läsnä kloroplastissa, on tärkeää, että synteesi koneisto ”on päällä” ja hajottava koneisto ”pois päältä” valossa.
Valo säätelee Calvinin syklin toimintaa • Muutokset strooman pH:ssa ja magnesium-ionien konsentraatiot ovat tärkeitä säätelijöitä. • Rubiscon aktivaatioon liittyy karbamaatti-magnesium kompleksin muodostuminen ja hiilidioksidi. • Strooman magnesium konsentraatio on myös tärkeä. Magnesiumin konsentraatio stroomassa lisääntyy 1-3 mM > 3-6 mM valossa. Kun valon vaikutuksesta protoneita kuljetetaan tylakoidikalvojen lumeniin niin magnesium-ioneja tuodaan stroomaan kompensoimaan positiivisten varausten menetystä. • Fruktoosi-1,6-bisfosfataasi ja vähäisemmässä määrin myös muut entsyymit aktivoituvat valossa seurauksena stroman pH:n muutoksiin.
Valoon liittyvät kovalenttiset muutokset säätelevät Calvinin syklin toimintaa • Pelkistynyttä ferredoksiinia tuotetaan fotosynteesin valoreaktioissa ja se reagoi hapettuneen tioredoksiini f tai m kanssa ferredoksiini-tioredoksiini-reduktaasi (FTR) entsyymin katalysoimana. Pelkistynyt tioredoksiini voi taas puolestaan pelkistää kohde-entsyymien disulfidi-siltoja. • Pimeässä entsyymien sulfhydryyli-ryhmät (SH) hapettuvat. • Eli entsyymit aktivoituvat pelkistyessään valossa ja deaktivoituvat hapettuessaan pimeässä. • Valon vaikutuksesta pelkistynyt tioredoksiini inhiboi katabolisia reaktioita (jotka tapahtuvat pääsääntöisesti pimeässä esim oksidatiivinen pentoosi fosfaatti reitti).
Rubisco voi toimia myös oksygenaasina • Ribuloosi-1,5-bisfosfaatti karboksylaasi / oksygenaasi • Substraattina on tällöin happi (eikä hiilidioksidi). • Hapen reagoidessa ribuloosi-1,5-bisfosfaatin (RuBP) kanssa syntyy 3-PGA ja 2-fosfoglykolaatti (2-hiilinen yhdiste). • Kaksi substraattia hiilidioksidi ja happi kilpailevat samasta entsyymin aktiivisesta kohdasta. Entsyymin aktiivisuus näiden substraattien suhteen riippuu paljon ympäristössä olevan hapen ja hiilidioksidin määrästä. Karboksylaatio tapahtuu kolme kertaa nopeammin kuin oksygenaatio, mutta ilmassa on 600 molekyyliä happea jokaista hiilidioksidi-molekyyliä kohden. Oksygenaasi aktiivisuudella onkin huomattavaa vaikutusta hiilidioksidin sitomistehoon. Joissakin tapauksissa (C3-kasvit) 50 % fotosynteesissä sidotusta hiilidioksidista menetetään fotorespiraation takia.
C3- kasvi C4- kasvi esimerkiksi ananas, maissi ja monet trooppiset kasvit
C4-fotosynteesi Esim. soijapavulla jopa 50 % sidotusta hiilestä menetetään fotorespiraation takia > C4 fotosynteesin kehittyminen eli Rubisco / Calvinin sykli kauemmaksi happea vapauttavasta fotosysteemistä
C3 / C4-kasvit C3- kasveilla valoreaktiot (happen vapautuminen) ja Calvinin sykli tapahtuvat samoissa mesofyllin soluissa > fotorespiraatio C4-kasveilla hiilidioksidi sidotaan mesofyllin soluissa ja kuljetetaan jännetupen soluihin, joissa Calvinin sykli tapahtuu. > ei vaaraa fotorespiraatiosta C4-kasvit toimivat paremmin korkeissa lämpötiloissa ja kuivilla mailla hirssipeltoa C4- kasvi http://omega.dawsoncollege.qc.ca/ray/Photo/phresp.htm
CAM = Crassulacean Acid Metabolism C4- kasveja muistuttava metaboliareitti, jossa CAM- reaktiot ovat ajallisesti erillään C3-reaktioista. Ilmaraot ovat yöllä auki ja päivällä kiinni. Yöllä hiilidioksidi liittyy PEP > oksaloetikkahappo > omenahappo, joka varastoituu vakuoliin. Päivällä omenahppo otetaan vakuolista, hiili- dioksidi vapautuu ja menee Calvinin sykliin.
C3, C4 ja CAM-kasvit http://www.mhhe.com/biosci/esp/2001_gbio/folder_structure/ce/m6/s5/