Download

1 / 32
330 likes | 543 Views
Macronutrients and plant nutrition. Peter Shaw. This version of the file contains very few pictures – less pretty but takes up MUCH less disc space and download time. To see the images in their full glory, make sure you come to the lecture!!. Overview.

E N D
Macronutrients and plant nutrition Peter Shaw This version of the file contains very few pictures – less pretty but takes up MUCH less disc space and download time. To see the images in their full glory, make sure you come to the lecture!!
Overview Today I want to look at all aspects of plant nutrient uptake. This will cover the plants’ viewpoint, but also soil conditions and management. Topics to cover: The biology of nutrient uptake by roots The soil chemistry affecting this uptake.
Root uptake It has long been appreciated that roots in plants are like guts in animals – the site where nutrients are taken up. Because of this plant roots usually have an immense surface area caused by repeated divisions. Much of this area is due to root hairs, and it is these which are the main sites of entry into a plant for water and nutrients. Root hairs adhere tightly to soil particles, which is where soil water tends to be bound. Water enters through the epidermis of root hairs into the apoplast of the root (extra-cellular space). Here is is gradually taken up by cells and enters the symplast, from where it passes the casparian strips into the xylem vessels of the stele.
The flow of water into roots is controlled by a band of corky, water-impermeable cells lining the root cortex which force water to flow into the main vessels symplastically. This band of corky tissue (suberin + lignin) is the casparian strip, and is present in the endodermis of the root systems of most vascular plants. The casparian strip ensures that all water entering the stele of the root (thence up to the main stem) has passed through a plasma membrane so has been regulated by transport proteins. Casparian strip stele Cortex
Note that the tracheids and vessel elements of the xylem are dead and lack protoplasts, hence their lumen is apoplast, not symplast. Minerals and water enter the xylem proper by being actively pumped from the walls of the endodermal (and stele parenchymal) cells. This way the xylem contents have been filtered through the plasma membranes of many cells, and are highly purified (of bacteria, mineral debris etc).
Mycorrhizas Root hairs also get infected by mycorrhizal fungi, which act as extensions of the root system and collect nutrients (especially P) for the plant. This is the normal state for most wild plants. In exchange the plant supplies sugars to the fungus. (This sugar drain can be a detectable cost, if nutrients are supplied as chemical fertilisers). The mycorrhizal condition has evolved at least 4 times, probably closer to 10, and can be over-simplified by shoehorning mycorrhizas into 2 groups: Sheathing (ecto) mycorrhizas, which envelop roots in a coat of hyphae, and endomycorrhizas which penetrate inside the cells inside a hosts’ root. In both cases the increase in uptake area is huge – as much as 3m of hyphae from 1cm root. There are many trials showing how much better plants do with mycorrhizal infection in poor soils.
VAM Or endomycorrhizas vesicles - these are large and stain well. Arbuscules are harder - really needing EM. It is not clear that all endos have these, casting doubt on the name VAM. Arbuscules die and are absorbed by the plant, only to have new arbuscules re-form. This seems to be how P is transferred to the plant.
TS of an ectomycorrhizally infected root Varieties of: What you actually see Sheath Hartig Net (between root cells)
The minerals • One of the classic stories in environmental science is that of Von Helmut’s willow. • He grew a willow tree in a pot for some years, adding only water. • During this time the plant gained weight, while the soil lost only c. half an ounce. He concluded that plants were composed of water (forgetting one key component: air) • It is true that given light, water + CO2 plants can grow for a while – but eventually growth will become stunted for lack of other nutrition.
All living things need certain vital elements • Analyse a plant, and you will find that it contains classes of biochemicals which require specific elements. Contains: CHO CHO CHO NS [CHO] NP [CHO] Mg [CHO] Fe [CHON] K, Ca [+lots else!] Class: Sugars fats carbohydrates proteins nucleic acids Chlorophyll cytochromes cytoplasm
Of these nutrients: • CHO are “free”, from air and water. • All the rest must be taken up (usually by roots from soil), and are vital nutrients. • The actual importance of these must be determined empirically by trials. • 150 years research has shown that crops routinely respond well to one set of elements – these are the macronutrients. • A 2nd set of elements don’t usually matter, but can cause specific deficiencies. These are the micronutrients. • Note that the relative importance of different elements is NOT indicated by their concentration in ashed material. Plants do not excrete wastes but accumulate them in old tissue, so that plant ashes contain metals such as aluminium which have no metabolic use.
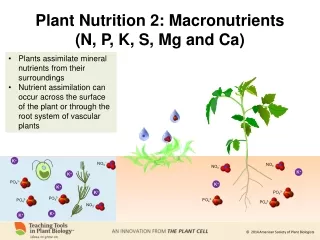
The macronutrients are: N, P, K [big 3] not forgetting Mg, S The micronutrients Ca, Fe, Mn, Mo, Na, Co, Si Crucially important point: It is not enough that a soil contains an element. What matters is its availability to plants. Thus acid digests of heathland soils show ample levels of nitrogen, but almost all of this is locked up in biomass or humic materials, and in fact such soils are acutely deficient in nitrogen.
Nitrogen • This is the most important plant nutrient, at least in terms of producing a reliable and large increase in growth. • Most of the advances in agricultural production since the 1940s have been won by vast increases in nitrogen application. • More N => more growth, typically soft fast sappy growth, deep green foliage. Our crops have been described as “nitrogen green”, referring to their force-fed status and colour. • Deficiency of N => stunting, yellowness and general poor growth.
Downside of N • N-rich plants are vulnerable to infection (pathogenic + saprotrophic fungi, sap suckers and herbivores all respond well to N). In part this explains the way that intensive farming is addicted to pesticides. • They are also mechanically softer so open to wind damage etc, and delay winter hardening so suffer more frost damage. • In Holland high levels of N pollution are causing forest dieback and heathland degeneration. This is a form of pollution called eutrophication (meaning “over-feeding”), leading to growth of weedy species at the expense of scarce forms. • Modern thatch lasts less well than victorian thatch, as modern plants experience higher levels of N in the environment.
N is chemically complex • It is everywhere, all the time, but in the useless form of an inert gas N2 • It also comes in 2 useful forms: ammonia NH3 ( ammonium NH4+), and nitrate NO3-. • These take part in a well known cycle, that you are strongly advised to learn:
N2 gas A simplified diagram of the nitrogen cycle After exposure to combustion flames Atmospheric NH3 NOx NO, N2O and N2 Atmosphere Soil Biological N Fixation Denitrification NO2- Nitrosomonas Nitrobacter Organic nitrogen and NH4+ Nitrification Release of H+ NO3- Biological reduction By nitrate reductase
Phosphorus, P • This is 2nd in effect after nitrogen. • Is always applied as phosphate PO43- • P deficiency leads to dark, stunted growth - often purple tints on the leaves, also causing poor root growth, so limited uptake of other nutrients as well. • P promotes cell division, mechanical strength, maturation/seed set, disease resistance. • Legumes need no N (are fixers) but do need P - has been suggested that a major conservation issue in UK is phosphate eutrophication, which leads to N enrichment.
Phosphate is tightly linked to the origins of the fertiliser industry • Traditionally the only source of phosphate was ground up bones and teeth (=forms of calcium phosphate). • William Buckland (notable Oxford Don) used guano to write “GUANO” on his college lawn. The letters stayed visible as deep green patches for years! • Richard Lawes in the 1840s discovered that bones dissolved in sulphuric acid made superb phosphatic fertiliser - is still used, called superphosphate. Bones + H2SO4 = £ • He used his £ to found Rothamsted experimental station in 1840s - still running trials 150 years later.
A feature of P: • Is that it is immensely immobile in soil. This is because many of its salts are highly insoluble, ie bones! Iron, aluminium phosphates are if anything even less soluble. • Fe3+ + PO43- => FePO4, insoluble • One practical consequence is that archaeological sites often show up as phosphate anomalies: P residues in tombs show where bones used to lie. Neolithic middens are detectable as P enriched. • P fertilisers once applied can be locked up in the soil without plants getting any benefit. Acid solutions (such as superphosphate) are the problem: acidity mobilises Fe, Al which them immobilise P. This process is confusingly called phosphorus fixation.
Managing Phosphate • The issue with P is not so much supply as availability. Pouring on P is no great help is the soil locks it up. (Some golf courses have such high levels of P that their soil could be sold as fertiliser under EU regulations!) Availability P in pure solution 4 5 pH 6 7 8 Availability Calcium level Fe, Al in soil solution 4 5 pH 6 7 8 Availability P in soil 4 5 pH 6 7 8
Managing phosphate 2 • This is achieved by controlling soil pH, which should be held around pH 6.5. • A prime function of liming acid soils is to elevate the pH to the 6-7 range. • Similarly, chalky soils may need to be acidified somewhat (perhaps as (NH4)2SO4) to increase P availability, though this is less usual. • Soils rich in Fe(OH)3 such as tropical clays will immobilise P whatever the pH
Potassium, K • Compared to the clear metabolic uses of P and N, the importance of K is rather surprising. Despite this, trials consistently show K to be a major determinant of plant growth. • It determines turgor, frost hardiness and resistance to wind. • K deficient plants are stunted and yellow - often the older needles / leaves are yellowest as this mobile element is translocated to the newest tissue. • Toadstools are remarkably high in K: 23% of the entire K in lawn soil may pass through the “fairy ring” toadstools Marasmius oreades
The problems with K.. • Are related to its mobility. • Left alone, K will rapidly leach out of soils or litter. It is low on acid sandy or organic soils, highest in clays. • Organisms take K up avidly, so that (uniquely among metals) its distribution is ecosystems is defined by biological not chemical processes. • K experiences luxury consumption - this means that plants take up more K than they need if it applied. It does them no harm, but does cost the farmer money. Best fertilise little but often.
Magnesium Mg2+ • This is one of the lime minerals, along with calcium. • It is needed in chlorophyll, so is vital for plant growth. • Its removal from acid forest soils leads to Type 1 German forest dieback, and magnesium deficiency can occur in UK crops. • In practice the UK rarely experiences much Mg deficiency as our rain is enriched with Atlantic seawater.
Magnesium deficiency.. • Causes yellowing, especially of older needles / leaves. • Is often caused by waterlogging - a physiological problem not a chemical one. • Is cured by adding dolomite (MgCO3+CaCO3) or purer magnesium salts.
Sulphur, as sulphate SO42- • This element is used in proteins due to the amino-acids cysteine and methionine. • Is prone to deficiencies on sandy soils low in OM, especially when put to grass or brassicas. (Old cabbage smells nasty because of the S in its oils). • Is a new problem - old fertilisers were so impure as to act as S supplements, and ambient SO2 levels used to be so high the S deficiency was unknown. • Now add as (NH4)2SO4 - but sparingly, as this is a strongly acidifying fertiliser.
Some micro-nutrients are of academic interest only – can be an issue in hydroponics with very pure chemicals, but not in real soils. • The ones that matter on a regular basis are: • calcium • Iron • Manganese • molybdenum • copper • zinc • boron
Calcium Ca2+ • Is a major ingredient of plant ashes and is a principal inter-cellular signalling element. • Hence one might expect it to be a major nutrient. In fact calcium deficiency is rare in crops, indeed virtually unknown for forests. • This is because of the link between Ca and soil pH – soils low in Ca are so acid that phosphorus is immobile (due to Fe3+/Al3+/Mn3+), which is the deficiency to show up first. • Ca problems are of course readily solved by addition of lime.
The commonest: Fe3+, Mn3+ • Iron and Manganese (don’t confuse Mn3+ with Magnesium Mg2+) are common soil elements used in metallo-enzymes (cytochromes). • In most plants, deficiencies of these elements is unusual. It occurs on chalky soils (pH>8) and is known as lime-induced chlorosis. It shows up as interveinal yellowing, with green veins on a yellow leaf.
The problem with Ericas: • There is one group of plants for which iron is a serious issue. • These are the ericaceous plants: heathers, rhododendrons, Camellias and allies. These are almost invariably plants of acid (pH<5) soils, where iron is always readily available. • These plants suffer when pH exceeds c. 5.5ish, due to iron’s immobility leading to deficiency. • Exactly why these plants need high levels of Fe is, I believe, unclear. The fact remains that they do, and garden centres never tell you this when you buy one!
A common tale of woe: • Is for an expensive new Rhododendron or Camellia to flower well in its first season in a new soil, then to lose colour, lose leaves, go yellow, sicken and die. • This is almost always due to the new soil being inadequately acid, hence iron deficiency. • Two solutions are on offer. • Plant it in a suitable soil!! (the only long-term solution) • Buy chelated iron (sequestrone etc). This contains EDTA = ethyl di-amine tetra-acetate, which reversibly locks up iron inside its molecule like a ball in a cage. Now the iron is isolated from water it dissolves whatever the pH, so is available to plant roots. Ericas are able to take up organically complexed iron, so stay alive a few more weeks..
EDTA – ethyldiamine tetra-acetate H H On paper O O O O C C C C N C C N C O O C C C O O H H How it actually works Fe