Download

1 / 20
250 likes | 606 Views
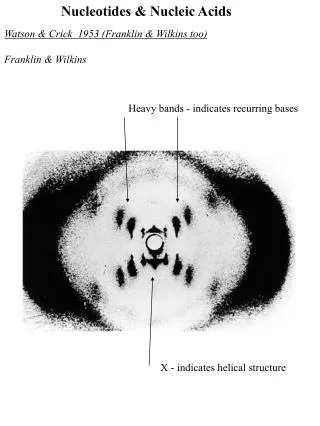
Chap. 8B Nucleotides and Nucleic Acids. Some Basics Nucleic Acid Structure Nucleic Acid Chemistry Other Functions of Nucleotides. Fig. 8-12. X-ray diffraction pattern of DNA fibers. Intro. to Nucleic Acid Chemistry.

E N D
Chap. 8B Nucleotides and Nucleic Acids • Some Basics • Nucleic Acid Structure • Nucleic Acid Chemistry • Other Functions of Nucleotides Fig. 8-12. X-ray diffraction pattern of DNA fibers.
Intro. to Nucleic Acid Chemistry The role of DNA as a repository of genetic information depends in part on its inherent stability. The chemical transformations that occur to DNA are generally very slow in the absence of an enzyme catalyst. However, even very slow reactions that alter DNA structure are physiologically significant. Processes such as carcinogenesis and aging are intimately linked to slowly accumulating, irreversible alterations of DNA. Other nondestructive alterations such as strand separation prior to DNA replication and transcription are essential to function. The chemical behavior of DNA is the focus of the next several slides.
Denaturation and Annealing of DNA Double-helical DNA can be denatured (melted) to single-stranded DNA by heating and extremes of pH. Disruption of the hydrogen bonds between paired bases and of base stacking causes unwinding of the double helix to form two single strands, completely separate from each other along the entire length or part of the length (partial denaturation) of the molecule (Fig. 8-26). Covalent bonds in the DNA are not broken by denaturation. When the temperature or pH is returned to the range in which most organisms live, the unwound segments of the two strands spontaneously rewind, or anneal, to yield the intact double helix. The renaturation of completed melted DNA occurs in two steps. First, the two strands slowly find each other by random collisions and form a short segment of complementary double helix. Second, the remaining unpaired bases rapidly zipper themselves together to form the complete double helix. The melting of double-helical DNA can be followed by measuring the increase in absorption of UV light (260 nm) on melting (the hyperchromic effect).
Heat Denaturation of DNA Every species of double-helical DNA has a characteristic denaturation temperature, or melting point (tm; formally the temperature at which half of the DNA is present as separated single strands) (Fig. 8-27). The melting point is dependent on, and rises with, the content of G/C base pairs in the DNA. This is because G/C base pairs are held together more tightly, by three hydrogen bonds, than are A/T pairs (two hydrogen bonds). The energetic requirements for DNA melting explain why DNA at replication origins, and at promoters used in gene transcription is enriched in A/T base pairs. RNA-RNA double helices and DNA-RNA hybrid double helices melt at higher temperatures than double-helical DNAs of comparable base composition, for unknown reasons.
DNA Hybridization The ability of two complementary DNA strands to pair (hybridize) with one another can be used to detect similar DNA sequences in two different species or within the genome of a single species (Fig. 8-29). To perform these analyses, the DNA samples to be compared are first completely denatured by heating. The solutions then are mixed and slowly cooled. Some DNA strands of each sample associate with their normal complementary partners and anneal to form duplexes. If the two DNAs have significant sequence similarity, they also tend to form partial duplexes or hybrids with each other. The greater the sequence similarity between the two DNAs, the greater the number of hybrids formed. The extent of hybrid formation reflects how closely related the organisms being analyzed are to one another. For example, human DNA hybridizes much more extensively with mouse DNA than with yeast DNA. Hybridization techniques are commonly used in many modern molecular biology procedures. (See Chap. 9).
Deamination of Nucleotides in DNA Purines and pyrimidines, along with the nucleotides of which they are a part, undergo spontaneous alterations in their covalent structure which can produce permanent changes (mutations) in the genetic information. One such modification is the spontaneous loss of the exocyclic amino groups (deamination) present in the bases of DNA (Fig. 8-30a). For example, deamination of cytosine in DNA to uracil occurs in about one of every 107 cytidine residues in 24 hours under cellular conditions. This corresponds to about 100 spontaneous events per day in a mammalian cell. This reaction likely explains why DNA contains thymine rather than uracil. Namely, uracils produced by cytosine deamination can be specifically recognized and repaired back to cytosine residues by enzymatic repair systems. Without repair, cytosine deamination would convert many G/C base pairs in DNA to A/U base pairs.
Depurination of Nucleotides in DNA Another important reaction in DNA is the hydrolysis of the N-ß-glycosyl bond between the base and the pentose, to create a DNA lesion called an AP (apurinic, apyrimidinic) site or abasic site (Fig. 8-30b). This reaction occurs at a higher rate for purines than for pyrimidines, and in the test tube is accelerated in the presence of dilute acid (pH 3). It is calculated that on the order of one in 105 purines (~10,000 per mammalian cell) are lost from DNA daily under cellular conditions. Again, repair systems must operate to repair abasic sites in DNA to prevent the accumulation of mutations.
Formation of Pyrimidine Dimers in DNA DNA also can be damaged by various forms of UV and ionizing radiation. For example, in the presence of near-UV light (200 to 400 nm), adjacent pyrimidine bases in nucleic acids combine via their rings to form cyclobutane pyrimidine dimers, and so-called 6-4 photoproduct pyrimidine dimers (Fig. 8-31a). Formation of a cyclobutane pyrimidine dimer introduces a bend or kink into the DNA (Fig. 8-31b). Pyrimidine dimers must be removed from the template strand for DNA replication to proceed normally. Higher-energy ionizing radiation, (x rays and gamma rays) can cause ring opening and fragmentation of bases as well as breaks in the covalent backbone of DNA. It is estimated that UV and ionizing radiations are responsible for about 10% of all DNA damage caused by environmental agents.
DNA-damaging Chemical Agents (I) DNA can be damaged by reactive chemicals introduced into the environment as products of industrial activity. Agents that result in deamination of bases are shown in Fig. 8-32a. All of these agents are precursors of nitrous acid (HNO2), which is the compound that actually is responsible for deamination. Bisulfate is also a deamination agent. Some of these chemicals are used in small amounts for food preservation.
DNA-damaging Chemical Agents (II) A broad class of chemicals that act as alkylating agents also cause a significant amount of damage to the bases of DNA (Fig. 8-32b). For example, dimethylsulfate ((CH3)2SO4) can methylate guanine to produce O6-methylguanine which can no longer base pair with cytosine. The compound S-adenosylmethionine is a cofactor used in enzymatic methylation of DNA. DNA methylation is important in bacterial restriction-modification systems and in mismatch repair of erroneously incorporated bases during replication. Probably the most important source of mutagenic alterations in DNA is oxidative damage.
Sanger DNA Sequencing (I) The Sanger method of DNA sequencing makes use of the mechanism of DNA synthesis by DNA polymerases (Fig. 8-33a). DNA polymerases require both a primer (a short oligonucleotide strand), to which nucleotides are added, and a template strand to guide the selection of each added nucleotide. The 3’-hydroxyl group of the primer reacts with an incoming deoxynucleoside triphosphate (dNTP) to form a new phosphodiester bond as the chain grows in the 5’ to 3’ direction.
Sanger DNA Sequencing (II) The Sanger method uses dideoxynucleoside triphosphate (ddNTP) analogs (Fig. 8-33b) to interrupt DNA synthesis. (The Sanger method is also known as the dideoxy or chain-termination method). When a ddNTP is inserted in place of a dNTP, strand elongation is halted after the analog is added, because the analog lacks the 3’-hydroxyl group needed for the addition of the next nucleotide. An overview of the steps performed in Sanger sequencing is presented in the next two slides.
Sanger DNA Sequencing (III) The DNA to be sequenced is used as the template strand, and a short oligonucleotide primer, radioactively or fluorescently labeled, is annealed to it (Fig. 8-33c). By addition of small amounts of a single ddNTP, for example, ddCTP, to an otherwise normal reaction system, the synthesized strands will be prematurely terminated at some locations where dC normally occurs. Given the excess of dCTP over ddCTP, the chance that the analog will be incorporated whenever a dC is to be added is small. However, ddCTP is present in sufficient amounts to ensure that each new strand has a high probability of acquiring a least one ddC at some point during synthesis. (Continued on the next slide).
Sanger DNA Sequencing (IV) The result is a solution containing a mixture of labeled fragments, each ending with a C residue. Each C residue in the sequence generates a set of fragments of a particular length, such that the different-sized fragments, separated by electrophoresis, reveal the location of C residues. This procedure is repeated separately for each of the four ddNTPs, and the sequence can be read directly from an autoradiogram of the gel. Because shorter DNA fragments migrate faster, the fragments located near the bottom of the gel represent the nucleotide positions closest to the primer (the 5’ end), and the sequence is read (in the 5’ to 3’ direction) from bottom to top. Note that the sequence obtained is that of the strand complementary to the strand being analyzed.
Sanger DNA Sequencing (V) Several high-throughput and automated sequencing methods, based on the Sanger method, are now used for rapid sequencing of large segments of DNA. One such method is illustrated in Fig. 8-34. In this approach, each of the four dideoxynucleotides used in chain-termination is labeled with a different fluorescent dye that gives all the fragments terminating in that nucleotide a particular color. All four labeled ddNTPs are added to a single reaction tube. The resulting dye-labeled segments of DNA copied from the template are applied to a single capillary gel and are subjected to electrophoresis. The DNA sequence is read by determining the sequence of colors in the peaks as they pass through a laser detector. Even more efficient methods for high-throughput sequencing are discussed in Chap. 9.
Chemical Synthesis of DNA DNA oligonucleotides of 70 to 80 nucleotides are routinely synthesized by the phosphoramidite method (Fig. 8-35). This automated method for DNA synthesis is conceptually similar to the synthesis of peptides on a solid support in that activating and protecting groups are attached to the reacting nucleotides to increase reaction specificity and yield.
Nucleoside Mono-, Di-, & Triphosphates The 5’ hydroxyl group of a nucleotide commonly may have one, two, or three phosphate groups attached to it. The resulting molecules are referred to as nucleoside mono-, di-, and triphosphates (Fig. 8-36). Starting from the sugar ring, the phosphates are labeled , ß, and . As discussed in the next slide, the hydrolysis of nucleoside triphosphates (particularly ATP) provides chemical energy needed to drive many cellular reactions. Nucleoside triphosphates also serve as the activated precursors of DNA and RNA synthesis.
ATP as a Source of Chemical Energy ATP is the nucleotide that is most commonly used as a source of energy for biological processes. The energy released by the hydrolysis of ATP (and the other nucleoside triphosphates) is accounted for by the structure of the triphosphate group. The bonds between the -ß and ß- phosphates of ATP are phosphoanhydride linkages. The hydrolysis of either of these bonds liberates about 30 kJ/mol under standard biochemical conditions (Fig. 8-37). When chemically coupled to an energy-requiring (endergonic) process, the hydrolysis of phosphoanhydride bonds often provides enough energy to drive the process forward. In contrast, the hydrolysis of the phosphoester linkage between the ribose and the phosphate of ATP is less exergonic, liberating about 14 kJ/mol.
Adenosine-containing Coenzymes A variety of enzyme cofactors serving a wide range of chemical functions contain adenosine (red shading) as part of their structure (Fig. 8-38). They are unrelated structurally except for the presence of adenosine, and in none of these cofactors does the adenosine moiety participate directly in the coenzyme function. Instead, it is recognized by the enzyme as an important “handle” in the binding of the coenzyme to the enzyme. The coenzymes shown in Fig. 8-38 play very important roles in metabolism. Coenzyme A functions in acyl group transfer reactions. NAD+ and FAD function in oxidation-reduction reactions.
Regulatory Nucleotides Hormonal signal transduction systems often rely on a nucleotide for intracellular signal transmission. These compounds (typically called second messengers) are formed by the binding of the hormone to a cell surface receptor, and cause changes in the activities of intracellular proteins and enzymes leading to the cellular response. Two common second messengers (cAMP and cGMP) are shown in Fig. 8-39. For example, cAMP plays a major role in epinephrine control of glycogen metabolism in the liver and skeletal muscle.