Download
1 / 59
590 likes | 815 Views
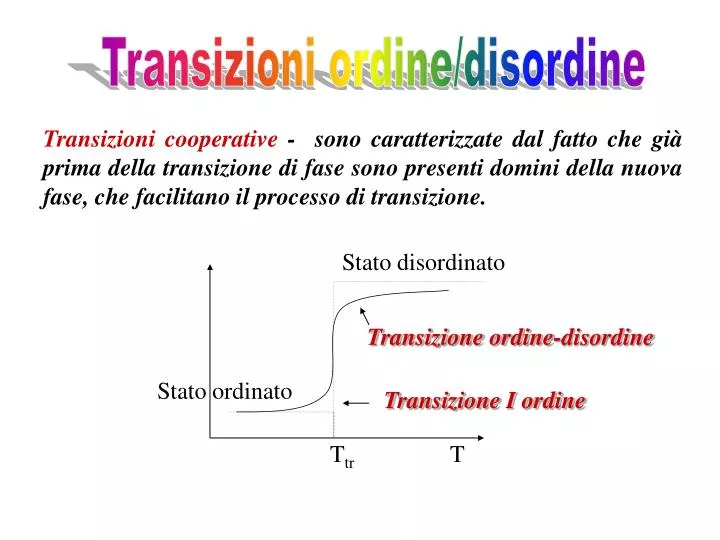
Transizioni ordine/disordine. Transizioni cooperative - sono caratterizzate dal fatto che già prima della transizione di fase sono presenti domini della nuova fase, che facilitano il processo di transizione. Stato disordinato. Transizione ordine-disordine. Stato ordinato.

E N D
Transizioni ordine/disordine Transizioni cooperative - sono caratterizzate dal fatto che già prima della transizione di fase sono presenti domini della nuova fase, che facilitano il processo di transizione. Stato disordinato Transizione ordine-disordine Stato ordinato Transizione I ordine Ttr T
Transizione elica-coil in polipeptidi gomitolo statistico ⇄ α-elica Tm = temperatura di transizione (m = melting, fusione) Frazione di α-elica: A T = Tm: fh = fc = 0.5
Transizione del I ordine L’andamento sigmoidale è tipico di una transizione cooperativa. La pendenza della curva a T=Tm è una misura della cooperatività della transizione.
Transizione coil-elica della poli(L-lisina) in funzione del pH Il pH di transizione (10.1) segue da vicino il pH di deprotonazione dei gruppi amminici (10.2): NH3+NH2 + H+
pH<10.2: il gruppo amminico laterale è protonato e la struttura elicoidale è destabilizzata dalle interazioni repulsive elettrostatiche tra le catene laterali. Lo stato di 'random coil' è invece stabilizzato da una solvatazione ottimale delle cariche elettrostatiche grazie alla maggiore libertà conformazionale del polipeptide. pH>10.2: predomina la forma deprotonata del gruppo amminico e le interazioni di legame idrogeno intramolecolare rendono più stabile la conformazione ordinata. Nel caso del poli(L-glutammico): COO- + H+COOH A pH acidi predomina la forma protonata e il polipeptide assume una struttura secondaria ordinata elicoidale, mentre a pH>4.5 predomina la forma deprotonata carica negativamente.
Transizione coil-elica del poli(L-glutammato) di sodio in funzione della percentuale di alcool isopropilico in miscele acqua/alcool.
Meccanismo della transizione coil-elica I step: Nucleazione Formazione del primo legame idrogeno i→i+4 Richiede l’immobilizzazione di 6 angoli torsionali (Ψ=-47°; Φ=-57°)
II step: Propagazione Formazione di un secondo legame idrogeno adiacente ad uno già formato. Richiede l’immobilizzazione di soli 2 angoli torsionali
Energetica della transizione Entalpia di solvatazione Formazione di legame idrogeno Interazione tra dipoli ammidici T=298K
Legame idrogeno vs. solvatazione Formazione di legame idrogeno in N-metil acetammide in diversi solventi
Competizione nella formazione di legame idrogeno: CCl4 < diossano < H2O
Entalpia di solvatazione Hsolv = differenza di solvatazione tra il peptide in elica e il peptide in coil. Un contributo generalmente sfavorevole alla transizione in elica (+2-3 kcalmol-1). Il coil è infatti libero di riaggiustarsi in modo da ottimizzare la solvatazione della sequenza peptidica (rigonfiamento).
Per un numero sufficientemente grande di residui (n10) il contributo attrattivo dovuto all’allineamento testa-coda dei dipoli predomina sul contributo repulsivo dei residui adiacenti, con una conseguente stabilizzazione della struttura elicoidale ( – 1-2 kcal·mol-1). L’effetto di questa interazione è quello di favorire lunghi tratti elicoidali.
Contributi entropici La valutazione dei contributi entropici del processo richiede una descrizione approfondita dei processi di solvatazione. Il contributo entropico alla transizione elicacoil è generalmente favorevole ed aumenta all’aumentare della temperatura:
Modello di Schellman Un modello a cooperatività infinita: le catene polipeptidiche possono esistere o in una configurazione tutta elica o in una configurazione tutto coil. fh=0.5 significa che il 50% delle catene è in elica e il 50% in coil. L’energia libera standard per la conversione di un singolo residuo da coil ad elica è: Un processo favorito entalpicamente, ma sfavorito dal punto di vista entropico.
Nella formazione di un α-elica 4 residui non possono formare legame idrogeno e i due terminali restano comunque liberi di ruotare: La variazione di energia libera dell’intero processo sarà:
Scriviamo il potenziale chimico dei due stati: All’equilibrio:
Dall’equazione di van’t Hoff: Poiché:
Modello Zimm-Bragg a) Modello per polipeptidi a basso peso molecolare Sono possibili solo configurazioni del tipo: cccccchhhhhhccccc o hhhhhhhcccccc Vi può essere cioè una sola regione di nucleazione per catena. Si trascurano effetti terminali di catena. Nel modello non vengono considerati i residui terminali, strutturalmente incapaci di dar luogo alla formazione di legami idrogeno.
Nel modello si identificano due contributi energetici: • Entropia di nucleazione: congelamento dei primi 4 angoli torsionali. • II. Propagazione: formazione di un legame idrogeno adiacente ad un residuo già in elica • …ccchcccc… ….ccchhcc….
I step: Nucleazione - formazione del primo legame idrogeno in una sequenza altrimenti in coil …cccccccc… ….ccchccc…. Consta di due contributi: <entropia di nucleazione> + <energia libera di propagazione> Il contributo dominante è quello entropico, il contributo entalpico può essere trascurato.
Dalla definizione statistica di entropia: Per un’elica perfetta: Per il gomitolo statistico si assume, applicando l’approssimazione degli isomeri rotazionali, che vi siano 3 stati popolati per ogni angolo torsionale (t, g+,g-):
Moltiplicando per i 4 angoli torsionali bloccati durante il processo di nucleazione: A 25°C:
Parametro di nucleazione (o di cooperatività) Per: Transizione non cooperativa! A T=300K: =2.5 kcalmol-1 0.001
Quanto minore è σ, tanto più grande è l’energia libera di nucleazione, tanto più cooperativa è la transizione.
Dalla definizione di : Snucl = Sh – Sc = RT ln Sh=0 (wh=1) Sc = - RT ln Tanto minore è il valore di , tanto maggiore sarà l’entropia del coil statistico, legata al numero di conformazioni accessibili al sistema, cioè al numero di isomeri rotazionali popolati. Nella serie glicina (R=H), alanina (R=CH3), leucina (R=CH2-CH(CH3)2), il valore di segue l’aumento dell’ingombro sterico della catena laterale (0.00001, 0.0008, 0.0033), proprio perché diminuisce il numero di disposizioni geometriche accessibili ad ogni angolo torsionale.
Parametro di propagazione Assume il significato di una vera e propria costante di equilibrio relativa al processo di propagazione delle’elica.
Gprop = - RT lns Nel caso dei residui glicina, alanina e leucina (s=0.60,1.06,1.14 rispettivamente) al crescere delle dimensioni della catena laterale, aumentano le interazioni attrattive di Van der Waals. Gruppi polari determinano l’instaurarsi di interazioni dipolari con il backbone peptidico, di cui bisogna valutare caso per caso l’entità sia rispetto al peptide in conformazione elicoidale, sia come gomitolo statistico. Nel caso dell’Asparagina (Asn, R=-CH2-CONH2), l’interazione della catena con il dipolo ammidico tende a destabilizzare l’elica.
Il parametro s è formalmente assimilabile ad una costante di equilibrio:
Per la formazione del primo legame idrogeno: Per la formazione del secondo legame idrogeno:
Per due residui adiacenti impegnati in legame idrogeno: Per tre residui adiacenti impegnati in legame idrogeno:
Per m residui adiacenti impegnati in legame idrogeno: Da cui si ottiene per la transizione in cui m residui passano da coil ad elica: Gtrans = Gnucl + m Gprop = - RT( ln + m ln s)
Dalla trattazione statistica dei vari modelli: Modello residui indipendenti (transizione non cooperativa)
Modello di Schellman (Cooperatività infinita) n = numero dei residui
Modello Zimm-Bragg b) Modello per polipeptidi ad alto peso molecolare In questo caso bisogna introdurre la possibilità di avere più centri di nucleazione. Per j centri di nucleazione e m residui adiacenti impegnati in legame idrogeno:
L’equilibrio tra sequenze lunghe e corte è in realtà il bilancio tra la tendenza a minimizzare l’energia e quella simultanea a massimizzare l’entropia. Per catene molto lunghe la tendenza, entalpicamente favorita, di formare lunghe sequenze ordinate è controbilanciata dalla perdita di entropia complessiva. Come risultato nella catena vi saranno piu centri di nucleazione con sequenze di eliche relativamente lunghe separate da regioni in coil (per esempio -turns in un filamento polipeptidico di una proteina globulare).
Il risultato per 0.8 < s < 1.2 è: Per s=1: Per s<1: Per s>1: per =110-4 s=0.8: fh=0.002 s=1.2: fh=0.997 per =110-2 s=0.8: fh=0.127 s=1.2: fh=0.837
Per =1 (Gnucl=0): Da cui si ottiene: In questo caso si ha completa assenza di cooperatività, tutti i residui si comportano in maniera indipendente e s coincide con la costante di equilibrio della transizione.
La cooperatività della transizione dipende da σ (più piccolo è σ, maggiore è l’energia libera di nucleazione, maggiore la cooperatività della transizione). Infatti:
A T=Tm, fh=0.5, s=1: Per σ→0: Per σ→1:
Un'eccezione significativa σ=2·10-4 n=1500 n=46 n=26 Poli(γ-benzil-L-glutammato) in dicloroacetico/dicloroetilene (80/20, v/v)