Download
1 / 30
550 likes | 3.13k Views
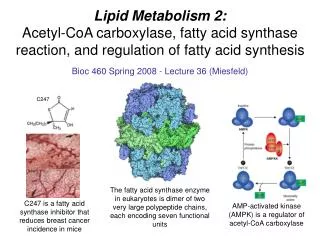
Lipid Metabolism 2: Acetyl-CoA carboxylase, fatty acid synthase reaction, and regulation of fatty acid synthesis. C247. Bioc 460 Spring 2008 - Lecture 36 (Miesfeld). The fatty acid synthase enzyme in eukaryotes is dimer of two very large polypeptide chains, each encoding seven functional units .

E N D
Lipid Metabolism 2:Acetyl-CoA carboxylase, fatty acid synthase reaction, and regulation of fatty acid synthesis C247 Bioc 460 Spring 2008 - Lecture 36 (Miesfeld) The fatty acid synthase enzyme in eukaryotes is dimer of two very large polypeptide chains, each encoding seven functional units C247 is a fatty acid synthase inhibitor that reduces breast cancer incidence in mice AMP-activated kinase (AMPK) is a regulator of acetyl-CoA carboxylase
Key Concepts in Lipid Metabolism • Fatty acid synthesis and degradation have several similarities and many differences. Both require carrier molecules and the enzymology of adding or subtracting acetate units to a hydrocarbon chain are similar. However, synthesis takes place in the cytosol, uses NADPH as coenzyme in redox reactions, and the building block is malonyl-CoA. • Acetyl-CoA carboxylase is the key regulated enzyme in fatty acid synthesis and is responsible for generating malonyl-CoA in a carboxylation reaction using acetyl-CoA. Acetyl-CoA carboxylase activity is regulated by both allostery (metabolic signaling) and phosphorylation (hormonal signaling). • The fatty acid synthase protein complex consists of six enzymatic activities and the acyl carrier protein (ACP). Seven reaction cycles are required to synthesize palmitate (C16) from 1 acetyl-CoA and 7 malonyl-CoA at cost of 14 NADPH. • The citrate shuttle is responsible for moving acetyl-CoA equivalents from the mitochondrial matrix to the cytosol when glucose levels are high and the citrate cycle is feedback inhibited by a high energy charge in the cell.
While the chemistry of the four core reactions required for the removal or addition of C2 acetyl groups to the hydrocarbon chain are similar between fatty acid degradation and synthesis, the two pathways are in fact quite distinct in terms of the required enzymes, subcellular location and source of redox energy. Fatty acid degradation occurs in the mitochondrial matrix and utilizes FAD and NAD+ as the oxidants in two oxidation reactions, whereas, fatty acid synthesis occurs in the cytosol and is dependent on NADPH serving as the reductant in the two corresponding reduction reactions. Other differences are listed below. Comparison of fatty acid synthesis and degradation
Review of Pathway Questions 1. What purpose does fatty acid synthesis serve in animals? • Fatty acid oxidation in mitochondria is responsible for providing energy to cells when glucose levels are low. Triacylglycerols stored in adipose tissue of most humans can supply energy to the body for ~3 months during starvation. • Fatty acid synthesis reactions in the cytosol of liver and adipose cells convert excess acetyl CoA that builds up in the mitochondrial matrix when glucose levels are high into fatty acids that can be stored or exported as triacylglycerols.
Review of Pathway Questions 2. What is the net reaction in the synthesis C16 palmitate? Fatty acid oxidation: Palmitate + 7 NAD+ + 7 FAD + 8 CoA + 7 H2O + ATP → 8 acetyl CoA + 7 NADH + 7 FADH2 + AMP + 2 Pi + 7 H+ Fatty acid synthesis:8 Acetyl CoA + 7 ATP + 14 NADPH + 14 H+→ Palmitate + 8 CoA + 7 ADP + 7 Pi + 14 NADP+ + 6 H2O
Review of Pathway Questions 3. What are the key enzymes in fatty acid synthesis?Acetyl CoA carboxylase - catalyzes the commitment step in fatty acid synthesis using a biotin-mediated reaction mechanism that carboxylates acetyl-CoA to form the C3 compound malonyl-CoA. The activity of acetyl CoA carboxylase is regulated by both reversible phosphorylation (the active conformation is dephosphorylated) and allosteric mechanisms (citrate binding stimulates activity, palmitoyl-CoA inhibits activity). Fatty acid synthase - this large multi-functional enzyme is responsible for catalyzing a series of reactions that sequentially adds C2 units to a growing fatty acid chain covalently attached to the enzyme complex. The mechanism involves the linking malonyl-CoA to an acyl carrier protein, followed by a decarboxylation and condensation reaction that extends the hydrocarbon chain.
Acetyl-CoA carboxylase catalyzes the commitment step in fatty acid synthesis which converts acetyl-CoA to malonyl-CoA Malonyl-CoA serves as the donor of C2 acetyl groups during each round of the fatty acid synthesis reaction cycle. The E. coli acetyl CoA carboxylase enzyme consists of three subunits which encode a biotin carboxylase, a biotin carrier protein and a transcarboxylase.
Acetyl-CoA carboxylase catalyzes the commitment step in fatty acid synthesis which converts acetyl-CoA to malonyl-CoA In the first step, the biotin carboxylase subunit of the enzyme uses ATP to form carboxyphosphate which is then dephosphorylated to drive the formation of carboxybiotin. The carboxybiotin arm then swings across the enzyme complex and positions the carboxyl group in a second active site where the transcarboxylase subunit transfers the carboxyl group from carboxybiotin to acetyl CoA to form the reaction product malonyl CoA.This same carboxyl group used to form malonyl CoA from acetyl CoA is removed by decarboxylation in step 4 of the fatty acid synthesis reaction cycle (decarboxylation is a highly exergonic reaction). Therefore, malonyl CoA essentially serves as the "activated" carboxylated form of acetyl CoA.
The fatty synthesis reaction cycle The four core reactions of fatty acid degradation and fatty acid synthesis are chemically similar although different enzymes are utilized and the two pathways are physically separated (degradation takes place in the mitochondrial matrix and fatty synthesis is a cytosolic pathway). Acetyl CoA enters the reaction cycle through malonyl CoA which is covalently linked to acyl carrier protein (ACP) through a thioester. Following decarboxylation of the malonyl group, and condensation with the enzyme-bound fatty acyl group, the extended hydrocarbon chain is chemically modified and then translocated from ACP back to the fatty acid synthase enzyme. The reduced ACP thiol is then ready to accept another malonyl group and start the cycle over again.
The fatty synthesis reaction cycle Acetyl-CoA is the priming group only in the first cycle, after that, only malonyl-CoA is added to the ACP carrier protein each time. There are four reaction steps required each cycle to result in the net addition two carbons to the growing fatty acid chain.
The fatty synthesis reaction cycle Each cycle of the fatty acid synthase reaction requires the input of one malonyl-CoA and the oxidation of 2 NADPH molecules (4 e- total). The synthesis of C16 palmitate therefore requires 14 NADPH.
The fatty synthesis reaction cycle In the final step, the enzyme palmitoyl thioesterase catalyzes a hydrolysis reaction to release palmitate.
The fatty synthesis reaction cycle Let us take a closer look at these reaction steps to see just how cool this fat making protein machine really is. In the first step of palmitate synthesis, an acetyl-CoA is used as a primer before the addition of the first malonyl-CoA. This "priming" reaction is mediated by the enzyme malonyl/acetyl CoA-ACP transacetylase (MAT) and only occurs during the first cycle of the reaction pathway. The sulfur atom in ACP is located at the end of a phosphopantetheine prosthetic group which is linked to a serine residue in the ACP protein. In step 2, the acetyl group attached to ACP is translocated to the thiol group of a cysteine residue in the -ketoacyl-ACP synthase (KS) subunit This translocation reaction is catalyzed by the KS enzyme itself and is required during each turn of the cycle.
The fatty synthesis reaction cycle Acetyl-CoA is added first to the ACP and then transferred to the KS subunit (not shown here).
The fatty synthesis reaction cycle The condensation reaction between the acetyl group on the KS subunit and the malonyl group on the ACP carrier protein is catalyzed by the -ketoacyl-ACP synthase (KS) subunit in which the acetyl group is transferred to malonyl-ACP in a decarboxylation reaction leading to the formation of acetoacetyl-ACP. Note that in subsequent cycles of the reaction, the growing fatty acyl chain is linked to the KS subunit and used in the condensation reaction with the malonyl group on ACP. In the next reaction, acetoacetyl-ACP is then converted to D-3-hydroxybutyryl-ACP through a reduction reaction catalyzed by -ketoacyl-ACP-reductase (KR) and NADPH oxidation. This is followed by a dehydration reaction catalyzed by -hydroxyacyl-ACP-dehydratase (DH) to form ,-trans-butenoyl-ACP, and a second NADPH-dependent reduction reaction catalyzed by the enzyme enoyl-ACP-reductase (ER) leading to the formation of butyryl-ACP. Lastly, the butyryl group is translocated to Cys163 of the KS subunit to regenerate ACP-SH which is then ready to accept another malonyl group in the next cycle.
Malonyl-CoA is always the incoming group and the condensation reaction, and subsequent modification reactions take place on the ACP carrier protein. In the last step (step 5 here), the extended chain is translocated to the KS subunit to make room for then next malonyl group. In the final reaction, palmitate is release from ACP by the enzyme palmitoyl thioesterase(TE) which hydrolyzes the fatty acid and regenerates the SH group on ACP.
Summary of the fatty synthesis pathway • We can now put the entire fatty acid synthesis pathway together by looking at the ATP and NADPH requirements for synthesizing one molecule of the C16 fatty acid palmitate from eight molecules of the C2 metabolite acetyl CoA. We begin by forming seven molecules of malonyl CoA using the acetyl CoA carboxylase reaction: • 7 Acetyl CoA + 7 CO2 + 7 ATP --> 7 malonyl + 7 ADP + 7 Pi • We then use these seven malonyl CoA molecules for seven turns of the reaction cycle beginning with the priming of fatty acid synthase by one molecule of acetyl CoA: • 1 Acetyl CoA + 7 malonyl CoA + 14 NADPH + 14 H+ --> • palmitate + 7 CO2 + 8 CoA + 14 NADP+ + 6 H2O There are 7 dehydration steps required for palmitate, why only 6 net H2O?
Summary of the fatty synthesis pathway • The net fatty acid synthesis reaction for palmitate (C18) can then be written as: • 8 Acetyl CoA + 7 ATP + 14 NADPH + 14 H+ --> • palmitate + 8 CoA + 7 ADP + 7 Pi + 14 NADP+ + 6 H2O • The 14 NADPH molecules required to synthesize one molecule of palmitate comes primarily from the pentose phosphate pathway (lecture 34), although some NADPH is also generated by reactions in the citrate shuttle as described in the next couple of slides. Write the net reaction for the synthesis of C18 stearate.
The citrate shuttle transports acetyl-CoA equivalents from the matrix to the cytosol and generates NADPH The majority of acetyl CoA used for fatty acid synthesis in the cytosol is derived from reactions that take place in the mitochondrial matrix. However, mitochondria do not contain an acetyl CoA transporter, therefore a shuttle system, called the citrate shuttle, is required to move the C2 units across the membrane. Citrate transport out of the mitochondria provides a mechanism to stimulate fatty acid synthesis in the cytosol when acetyl CoA accumulates in the mitochondrial matrix. This build-up of acetyl CoA occurs when high glucose levels stimulate the conversion of pyruvate to acetyl CoA resulting in a high energy charge in the cell and feedback inhibition of the citrate cycle reactions. Under these conditions, citrate synthase produces citrate from acetyl CoA and oxaloacetate which is then transported to the cytosol rather than being converted to isocitrate by the enzyme aconitase.
The citrate shuttle transports acetyl-CoA equivalents from the matrix to the cytosol and generates NADPH Once in the cytosol, the citrate is cleaved by the enzyme citrate lyase to generate cytosolic acetyl CoA and oxaloacetate. The acetyl CoA is used for fatty acid synthesis and the oxaloacetate is converted to malate by cytosolic malate dehydrogenase. The production of cytosolic NADPH by malic enzyme provides additional reducing equivalents for fatty acid synthesis and supplements the NADPH generated by the pentose phosphate pathway.
Regulation of fatty acid synthesis The primary control point for regulating flux through the fatty acid biosynthetic pathway is the modulating the activity of acetyl CoA carboxylase. The activity of acetyl CoA carboxylase is controlled by both allosteric mechanisms (metabolic control) and covalent modification (hormonal control). Acetyl CoA carboxylase is most active when it is in a homopolymeric form. Citrate and palmitoyl CoA are metabolites that bind to an allosteric site on the enzyme stimulating polymerization or depolymerization, respectively.
Metabolic regulation of acetyl-CoA carboxylase Allosteric regulation of acetyl CoA carboxylase activity makes sense because when cytosolic citrate levels are high it means that the citrate shuttle is active and fatty acid synthesis is favored. However, when palmitoyl-CoA levels in the cytosol are high, it serves as a feedback inhibitor to decrease flux through the fatty acid synthesis pathway.
Hormonal regulation of acetyl-CoA carboxylase Hormone signaling also regulates the activity of acetyl CoA carboxylase. Insulin signaling leads to dephosphorylation and enzyme activation (polymerization), whereas, glucagon signaling results in phosphorylation and enzyme inactivation (monomeric form).
Regulation of acetyl-CoA carboxylase activity Insulin activates acetyl CoA carboxylase activity by stimulating dephosphorylation through protein phosphatase 2A (PP2A). In contrast, glucagon and epinephrine signaling activate the enzyme AMP-activated protein kinase (AMPK) which phosphorylates acetyl CoA carboxylase and shifts the equilibrium to the inactive monomeric form. Insulin signaling is activated by high serum glucose levels, and therefore activation of acetyl CoA carboxylase activity ensures that excess glucose will be rapidly converted to fatty acid for long term energy storage. Similarly, glucagon or epinephrine signaling is activated by low serum glucose levels, or neuronal input, respectively, and they lead to inhibition of acetyl CoA carboxylase activity to spare glucose for other purposes. Importantly, citrate binding to phosphorylated acetyl CoA carboxylase can result in partial enzyme activation by shifting the equilibrium in favor of polymer formation. This mechanism provides a way for the cell to respond to short term metabolic changes (excess citrate) by stimulating fatty acid synthesis even before long term hormone signaling is activated.
AMPK is an important metabolic sensor The regulatory protein AMPK is activated by low energy charge in the cell (high levels of AMP). The activity of AMPK is regulated by both AMP binding and by phosphorylation at a highly conserved threonine residue. The enzyme that phosphorylates AMP kinase is functionally referred to as AMP kinase kinase (AMPKK). When the energy charge in the cell is low, then AMPKK activity is stimulated by AMP binding, leading to activation of AMPK and inhibition of acetyl CoA carboxylase. However, when glucose levels are high, insulin signaling stimulates the activity of protein phosphatase 2C (PP2C), resulting in dephosphorylation of AMPK and accumulation of active acetyl-CoA carboxylase. The net result is an increase in fatty acid acid synthesis which makes sense because when glucose levels are high, it is important to store stimulate fatty acid synthesis.
Three Metabolic Control Points of FA Synthesis There are three metabolic control mechanisms that regulate flux through the fatty acid synthesis pathway. Excess acetyl CoA in the mitochondria results in citrate export to the cytosol which activates acetyl CoA carboxylase activity (stimulates enzyme polymerization), thereby producing malonyl CoA. Malonyl CoA inhibits carnitine acyltransferase I activity to prevent mitochondrial import and degradation of newly synthesized fatty acyl CoA molecules. When palmitoyl CoA levels exceed the metabolic needs of the cell, feedback inhibition of acetyl CoA carboxylase activity by palmitoyl CoA (stimulates enzyme depolymerization) decreases flux through the fatty acid synthesis pathway.
Three Metabolic Control Points of FA Synthesis What is the likely metabolic fate of the palmitoyl-CoA if this were a liver cell? What if it were a fat cell?