Download
1 / 50
540 likes | 800 Views
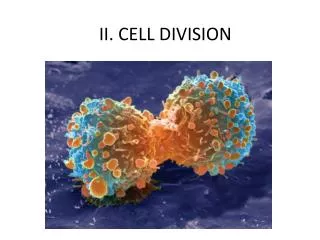
Apicomplexan host cell invasion II. Different parasites use different mechanisms to invade cells. Trypansoma cruzi -- recruits endosomes and later escapes into the cytoplasm Leishmania -- induces phagocytosis and thrives in a mature lysosomal compartment

E N D
Different parasites use different mechanisms to invade cells • Trypansoma cruzi -- recruits endosomes and later escapes into the cytoplasm • Leishmania -- induces phagocytosis and thrives in a mature lysosomal compartment • Mycobacterium tuberculosis -- induces phagocytosis and blocks lysosomal maturation • Toxoplasma -- resides in a specialized parasitophorous vacuole, how does it get in there and does this involve phagocytosis by the host cell?
The parasitophorous vacuole is not fusing with lysosomes • Macrophages were incubated with life (A/B) and heat killed (C/D) parasites • Note that only vacuoles containing heat killed parasites show staining for a lysosomal marker protein. • Dead parasites go in by phagocytosis, living parasites enter differently Joiner et al., Science 249:641-6
Invasion depends on parasite not host cell actin Salmonella • Host cell invasion by Salmonella and Toxoplasma can be inhibited by cytochalasin D, a drug that prevents actin polymerization • In Salmonella, which is taken up by phagocytosis, invasion can be rescued by using a drug resistant host cell mutant (Cyt1) • Toxoplasma invasion remains inhibited in the drug resistant host cell. • (black bars drug resistant, white bars drug sensitive host cell) • Generating a drug resistant parasite rescues invasion Toxoplasma Dobrowolski JM, Sibley LD. Cell. 1996 84(6):933-9.
Invasion depends on parasite not host cell actin • Cytochalasin treatment does not to appear to inhibit attachment (bar graph in c shows number of parasites bound to cells at different drug doses) • CytD inhibits the movement of the parasite into the host cell. • A parasitophorous vacuole (PV) is still set up, however the parasite can not move in, and the moving junction remains at the apical tip of the parasite Dobrowolski JM, Sibley LD. Cell. 1996 84(6):933-9.
Apicomplexan host cell invasion Movies Dr. Gary Ward, Univ. of Vermont http://www.uvm.edu/~mmg1/videos_ward.php?id=23
Invasion depends on sequential protein secretion from three organelles • Micronemes (Mn): secretion of micronemes brings protein to the parasite surface that provide ‘traction’ • Microneme proteins are required for gliding & invasion, the cleanest example is the Plasmodium sporozoite protein TRAP (Mic2 is a T. gondii homolog that is functionally equivalent in the tachyzoite) • Microneme proteins can bind to a variety of carbohydrates found on the surface of cells and other biological materials • Microneme proteins are assembled into complexes in the ER, mature by proteolysis, are stored in micronemes and are secreted at the apical tip of the parasite upon stimmulation • How does this help moving and how are they linked to the internal actin-based engine? Sultan et al, Cell 90: 511-522
The conveyor-belt model of gliding motility • Gregarines are a group of ‘primitive’ apicomplexans which parasitize invertebrates • In comparison ot Toxoplasma or the malaria parasite these are fairly large cells which makes them easier to study Movies by Dr. C. King (University College, London)
The conveyor-belt model of gliding motility • Beads attached to the surface of gregarines are ‘treadmilled’ to the end of the parasite cells • This suggests that the gliding machine moves microneme proteins over the surface from the apical to the basal end of the parasite
Gliding parasites deposit proteins and lipid trails • Similar to a slug the parasite leaves behind a trail of surface proteins and lipids • A protease cleaves microneme proteins at the end of the cell to detach and allow for propulsion (sheddase) • What is the force driving microneme proteins -- actin polymerisation or an actin/myosin motor?
A special myosin is required to move microneme proteins over the surface Normal myosin • Toxoplasma and other Apicomplexa have a parasite specific myosin (MyoA) • This myosin is localized right under the surface membrane of the parasite • Using genetic engineering a mutant parasite was constructed in which this myosin can be suppressed • Suppression of myosin result in loss of parasite motility (as seen for actin loss of motility also causes loss of host cell invasion) • What are the “gears” that connect this motor to the Mic tires and which part is anchored in the parasite? Suppressed myosin Meissner et al., Science. 298:837-40
PM IMC MT The gliding machinery is anchored in the inner membrane complex
The gliding machinery is anchored in the inner membrane complex • Biochemical studies aimed at identifying pellicle proteins let to the discovery of GAP45 a protein associated with outer face of the IMC • GAP45 was associated with the IMC despite the fact that is primary sequence suggested that this should be a soluble protein http://www.jcb.org/cgi/content/full/165/3/383
GAP45 is part of a complex including GAP50, MyoA & MLC http://www.jcb.org/cgi/content/full/165/3/383
The treadmilling model of apicomplexan gliding motility Soldati et al., Trends in Parasitology 20: 567-574
The conveyor-belt model • Motility depends of parasite actin/myosin (MyoA) • Myosin is anchored into the outer IMC membrane (GAP45/50, MyoA, MLC) • Short actin filaments form and are moved towards the posterior end of the parasite by the myosin power stroke • The short actin filaments are linked to microneme proteins by an adaptor (aldolase) -- movement of actin filaments results in movement of microneme proteins • Microneme proteins are shed at the back end (rhomboid proteases are the best candidates for this activity) • The parasite glides over the substrate
Invasion of the red blood cell by the malaria parasite • Although there are significant differences in the host cells which are invaded by different apicomplexa the mechanism seems conserved • Here the Plasmodium merozoite/red blood cell example
Invasion of the red blood cell by the malaria parasite • A moving junction is formed tightly opposing parasite and host cell membrane and separating parasitophorous vacuole lumen and outside medium
Secretion of the rhoptries is associated with PV formation • Rhoptries (Rh): secretion of rhoptries is required for the formation of the parasitophorous vacuole • Like micronemes rhoptries are secreted at the apical tip of the parasite • Some rhoptry proteins make up (part) of the moving junction (they are stored in the neck portion of the organelles and called RONs) • Other rhoptry proteins a found throughout the membrane of the parasitophorous vacuole after secretion and are stored in the bulbous part (ROPs) • A third group of very interesting rhoptry proteins is injected into the host cell and manipulates gene expression in the host nucleus -- ‘kiss and spit’ (check out review on the class web site)
Kiss & spit (thank John Boothroyd for the term) • It appears that a variety of rhoptry proteins are directly injected into the host cell and that this is involved in the formation of the parasitophorous vacuole • Note that staining for the rhoptry protein ROP1 highlights the parasitophorous vacuole (arrow) as well as numerous vesicles (arrowheads) formed at the invasion site within the host cell cytoplasm • Does the parasite “inject” the PV membrane?
Where does the membrane for the parasitophorous vacuole come from? • The PV membrane is derived from the host cell plasma membrane • The PV is provided by the parasite (e.g. by secretion from the rhoptries) • Both contribute to the PV
Where does the membrane for the PV come from? • Patch clamp cells and follow invasion by video microscopy • Certain electric properties of the cell (their capacitance) can be used as a measure of their total surface membrane • If membrane is parasite derived the surface area should grow during invasion if it is derived from the host cell surface the area should stay constant
There is no significant change of host cell surface during invasion
Host cell surface area decreases after PV pinches off • Cell surface area remains constant over invasion • When the parasitophorous vacuole pinches off the cell surface area drops • The parasitophorous vacuole is derived from the membrane of the host cell
Rhoptry proteins organize the moving junction • The parasite protein Ron4 (red) localizes exactly to the moving junction during invasion • Note that other parasite surface proteins (green) do not enter the parasitophorous vacuole but are shed at the moving junction • In non invading parasites Ron4 is stored in the rhoptry (necks, green, compare to micronemes red)
Rhoptry proteins have multiple functions in vacuole formation • Several RON proteins (2/4/5) assemble into a complex in the rhoptry neck • The complex is secreted and inserts into the host membrane (this is not fully defined yet but might parallel E. coli’s insertion of its own receptor) • AMA1 is a special microneme protein that engages the RON complex and serves as a specialized invasion ligant • ROPs are secreted into the cytoplasm and fuse back to the PV
The moving junction skims proteins out of the membrane as the PV forms • The surface membrane of the host cell was labeled for protein (green) and lipid (red) prior to infection with Toxoplasma (blue) • Note that while the lipids are clearly visible in the vacuole the proteins are excluded
Rhoptry proteins modify the function of the host cell nucleus • Several rhoptry proteins are injected into the host cell cytoplasm during invasion • They accumulate in the host cell nucleus • Interestingly, many of them are enzymes capable of changing the phosphorylation state of proteins (kinases & phosphatases) • Their precise function remains to be determined but it appears that they modulate gene expression in the host cell and that their activity is required for rapid growth and the ability to cause disease (virulence)
Rhoptry proteins have multiple functions in vacuole formation & host manipulation Bradley & Sibley, Current Opinion in Microbiology 10: 582-587
Invasion depends on sequential protein secretion from three organelles • Dense granules (DG): secretion of dense granules occurs after the vacuole is fully formed and continues throughout the intracellular growth of the parasites • Dense granules are secreted from the basal end of the parasites • Dense granule proteins likely play a role in modification of the vacuole into an environment supportive of parasite growth
Dense granule proteins are secreted into the PV • Certain dense granule proteins are soluble in the lumen of the PV others integrate into the membrane • These proteins are probably involved in modifying the vacuole
The PV is highly modified to suite the parasite’s needs • Tubular network increases surface (dense granule) • Sieving pores give access to small nutrient molecules in the host cell cytoplasm (probably dense granule) • Specific host cell organelles are recruited close to the PV membrane (rhoptry)
Dense granules are involved in establishing the intravacuolar network
Apicomplexan invasion • Active, parasite driven process • Depends on parasite actin/myosin motility (conveyor belt model) • Involves secretion of micronemes (attachment, motility), rhoptries (PV & MJ formation) and dense granules (makes PV into a suitable home) • Sets up a parasitophorous vacuole which initially is derived from the host cell cell-membrane • A moving junction is formed which screens out host membrane proteins from the PV, the PV is fusion incompetent and the parasite protected
Microneme protein complexes interact with the the host cell • Micronemes contain a large set of proteins containing protein and carbohydrate binding and interaction domains • MIC proteins associate into complexes in the ER and this association is critical for protein targeting • Proteolytic processing is critical for complex association, maturation and finally shedding Dowse T et al. Host cell invasion by the api...[PMID: 15358257]
Microneme protein complexes interact with the the host cell • The different roles of individual microneme proteins has been studied using conditional KOs in Toxoplasma (also see TRAP study by Sultan in Plasmodium) • MIC 2 is required for gliding and invasion (no ‘full’ block of invasion) • AMA1 is critical for invasion yet dispensable for gliding motility • This could suggest different ‘sets of tires’ for locomotory motility and invasion motility -- both depending on the actin/myosin engine http://www.molbiolcell.org/cgi/content/full/16/9/4341 doi:10.1371/journal.ppat.0020084
How to let go - rhomboid proteases in Toxoplasma & Plasmodium • Sequencing of ‘shed’ soluble adhesins suggest cleavage within the transmembrane region • Most characterized apicomplexan adhesins show conserved amino acids in this region which are similar to those demonstrated to be critical for rhomboid proteases in Drosophila • Rhomboids are transmembrane proteins and act as proteases cleaving the target with in the TM region http://www.pnas.org/cgi/content/full/102/11/4146 http://www.jcb.org/cgi/content/full/174/7/1023 Dowse TJ et al. Rhomboid-like proteins in Api...[PMID: 15922242]
How to let go - rhomboid proteases in Toxoplasma & Plasmodium • Several rhomboids are encoded by apicomplexan genomes and they are differentially expressed over the life cycle • TgROM5 in T. gondii localizes to the surface, it’s distribution is patchy and often at the posterior in gliding parasites. • TgROM5 (but not other ROMs) is capable of shedding MIC2 • Similar work in Plasmodium confirms the rhomboid model http://www.pnas.org/cgi/content/full/102/11/4146 http://www.jcb.org/cgi/content/full/174/7/1023 Dowse TJ et al. Rhomboid-like proteins in Api...[PMID: 15922242]