Download

1 / 72
1.19k likes | 3.55k Views
Lehninger Principles of Biochemistry, Fourth Edition, 2005. Chapter 4 THE THREE-DIMENSIONAL STRUCTURE OF PROTEINS. 中央研究院 生物化學研究所 曾湘文 博士 Mar. 13, 2007. Outlines:. 4.1 Overview of Protein Structure 4.2 Protein Secondary Structure 4.3 Protein Tertiary and Quaternary Structures

E N D
Lehninger Principles of Biochemistry, Fourth Edition, 2005 Chapter 4THE THREE-DIMENSIONALSTRUCTURE OF PROTEINS 中央研究院 生物化學研究所 曾湘文 博士 Mar. 13, 2007
Outlines: 4.1 Overview of Protein Structure 4.2 Protein Secondary Structure 4.3 Protein Tertiary and Quaternary Structures 4.4 Protein Denaturation and Folding
Levels of Structure in Protein • Primary: A description of all covalent bonds. The sequence of AA residues • Secondary: particularly stable arrangements of AA giving rise to recurring structural patterns. • Tertiary: All aspects of the 3D folding of a polypeptide. • Quaternary: The spatial arrangement of multisubunits protein
The structures of proteins determine their functions • A description of all covalent bonds (mainly peptide bonds and disulfide bonds) linking amino acid residues in a polypeptide chain is its primary structure. The most important element of primary structure is the sequenceof amino acid residues. • Secondary structurerefers to particularly stable arrangements of amino acid residues giving rise to recurring structural patterns. • Tertiary structuredescribes all aspects of the three-dimensional folding of a polypeptide. • When a protein has two or more polypeptide subunits, their arrangement in space is referred to as quaternary structure.
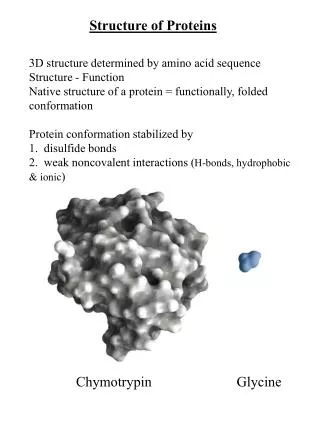
Three-dimensional structure of protein • The three-dimensional structure of a protein is determined by its amino acid sequence. • The function of a protein is determined by its structure. • An isolated protein has a unique structure. • The most important forces stabilizing the specific structure maintained by a given protein are noncovalent interactions. • Similar structure of proteins assists our understanding of protein architecture.
4.1 Overview of Protein Structure • The conformations existing under a given set of conditions are usually the ones that are thermodynamically the most stable, having the lowest Gibbs free energy (G). • Proteins in any of their functional, folded conformations are called native proteins.
A protein’s conformation is stabilized Largely by Weak Interactions • Proteins in any of their fictional, folded conformations are called native proteins • Stability: the tendency to maintain a nativeconformation. • The protein conformation with the lowest energy (i.e., the most stable conformation) is the one with the maximum number of weak interaction. • Hydrophobic AA side chains tend to be clustered in a protein’s interior, away from water. • Hydrogen bonds between groups in protein form cooperatively. Formation of one hydrogen bond facilitates the formation of additional bonds • Simple rules: • hydrophobic residues are largely buried in the protein interior; • the number of hydrogen bonds within the protein is maximized.
Three bonds separate sequential a carbons in a polypeptide chain The N-Caand Ca -C bonds can rotate, with bond angles designated φ(phi) andψ (psi), respectively. The peptide CON bond is not free to rotate. Other single bonds in the backbone may also be rotationally hindered, depending on the size and charge of the R groups. In the conformation shown, φ and ψ are 180℃ (or - 180℃). As one looks out from the carbon, the φ and ψ angles increase as the carbonyl or amide nitrogens (respectively) rotate clockwise.
The planar peptide group and Ramachandran plot for L-Ala residues 0 • The bond angles resulting from rotations at Ca for N-Ca - F (phi)and Ca-C - y (psi)are both defined as 0o when the two peptide bonds flaking that a carbon are in the same plane and positioned as shown. • Both f and y are defined as 180o when the polypeptide is in its fully extended conformation and all peptide groups are in the same plane Ramachandran plot
Ramachandran plot alpha-helix beta-strand
4.2 Protein Secondary Structure • The αHelix Is a Common Protein Secondary Structure • Amino Acid Sequence Affects Helix Stability • The βConformation Organizes Polypeptide Chains into Sheets • βTurns Are Common in Proteins • Common Secondary Structures Have Characteristic Bond Angles and Amino Acid Content
a helix • The polypeptide backbone is wound around an axis, and the R groups of the aa protrude outward from the helical backbone. Each helical turn includes 3.6 AA. • The amino acid residues in an a helix have conformations with y -45o to -50o and F -60o. • A single turn extends about 5.4 A along the axis, • The helix found in all protein is right-handed, about ¼ of all aa in peptides are found in a helices. • The a helix is stabilized by a hydrogen bond between the H atom attached to the electronegative nitrogen atom of a peptide and the electronegative carbonyl oxygen atom of the fourth aa on the amino-terminal side of that peptide bond- intra H bond • Each successive turn of the ahelix is held to adjacent turns by three to four hydrogen bonds
an helix can form in polypeptides consisting of either L- or D-amino acids. However, all residues must be of one stereoisomeric series; a D-amino acid will disrupt a regular structure consisting of L-amino acids, and vice versa. Naturally occurring L-amino acids can form either right- or left-handed helices, but extended left handed helices have not been observed in proteins. Right-handed vs. Left handed
Interactions between R groups of Amino acids three residues apart in an a helix • The polypeptide backbone (carbons, -amino nitrogens, and -carbonyl oxygens) is shown in gray for a helix segment 13 residues long. The only side chains represented here are the interacting Asp100 (red) and Arg103 (blue) side chains. • Two aromatic aa are often similarly spaced--- hydrophobic interaction. • Pro---introduces a destablizing kink , the N atom has no substituent H to participate in H bonds. • Glycine occurs infrequently in a helices- more conformational flexibility - Polymers of glycine tend to take up coiled structures quite different from an ahelix.
The electric dipole of a peptide bond- Helix dipole • A small electric dipole exists in each peptide bond. These dipoles are connected through the hydrogen bonds of the helix, resulting in a net dipole extending along the helix that increases with helix length • Negatively charged aa (N terminal); Positively charged aa (C terminal). will stabilize a helix
Five constraints affect the stability of an a helix • Electrostatic repulsion (or attraction) • Bulkiness of adjacent R groups • The interactions between aa side chains spaced three (or four) residues apart. • The occurrence of Pro and Gly. • The interaction between aa at the ends of the helix and the electric dipole inherent to the a helix.
The b conformation - b sheet Fully extended f, y = 180o
b conformation: b sheet • The backbone of the polypeptide chain is extended into a zigzag, and arranged side-by-side to form a structure resembling a series of pleats. • H-bonds are formed between adjacent segments---inter H bonds • The peptide chains can be either parallel or antiparallel. The structures are somewhat similar, although the repeat period is shorter for the parallel conformation (6.5 Å, versus 7 Å for antiparallel) and the hydrogen bonding patterns are different. • R groups of the aa on the touching surfaces must be relatively small--- Gly, Ala.
b turns and the trans- and cis- isomers of a peptide • 1/3 of aa are in turn or loop in globular proteins. • b turns: 4 aa; 180o turn, The carbonyl oxygen of the first aa forming a H bond with amino-group H of the fourth. • The central two aa do not have inter-residue H bonding--- H bonding with water. • Gly--- small and flexible; Pro--- peptide bonds involving the imino nitrogen of Pro assume the cis configuration- tight turn. • g turn, a 3 aa turn with a H bond between the first and third aa.
Ramachandran plots for a variety of structures • Every type of secondary structure can be completely described by the bond angles and at each residue. • Most values of Fand ytaken from known protein structures fall into the expected regions, with high concentrations near the a helix and bconformation values as predicted • Glycine is often found in a conformation outside these regions - Because its side chain, a single hydrogen atom,is small .
Relative probabilities that a given amino acid will occur in the 2nd structure • Some amino acids are accommodated better than others in the different types of secondary structures. • Ex. common presence of Pro and Gly residues in turns and their relative absence in ahelices, are readily explained by the known constraints on the different secondary structures.
4.3 Protein Tertiary and Quaternary Structures • Fibrous Proteins Are Adapted for a Structural Function • Structural Diversity Reflects Functional Diversity in Globular Proteins • Myoglobin Provided Early Clues about the Complexity of Globular Protein Structure • Globular Proteins Have a Variety of Tertiary Structures • Protein Motifs Are the Basis for Protein Structural Classification • Protein Quaternary Structures Range from Simple Dimers to Large Complexes
Protein tertiary and Quaternary structures • The overall three-dimensional arrangement of all atoms in a protein is referred to as the protein’s tertiary structure. • The arrangement of protein subunits in three-dimensional complexes constitute quaternary structure. • Fibrous protein: peptide chains arranged in long strands or sheets.--- a single type of secondary structure--- adapted for a structural function. • Globular protein: peptide chains folded into a spherical or globular shape.--- several type of secondary structure.--- enzyme and regulatory protein.
Fibrous Proteins Are Adapted for a Structural Function • a-Keratin, collagen, and fibroin (silk) are fibrous proteins commonly found in animals. • Their structural simplicity makes them a useful place to begin our investigation of protein tertiary
a-keratin • Found in mammals: hair, wood, nail, claw, quills, horns, hooves… outer layer of skin. • One of intermediated filament protein (most in the cytoskeletons). • The a keratin helix is right-handed a helix. Two keratin helix form a left-handed coiled coil ---supertwist. • 2 a helices --- hydrophobic AA, meshed together in a regular interlocking pattern---closely packing--- Ala, Val, Leu, Ile, Met, and Phe are rich. • The cross-links stabilizing quaternary structure are disulfide bond. • The hardest and toughest α-keratins, such as rhinoceros horn, up to 18% of the residuesare cysteines.
Disulfide bonds and permanent waving hair • Hair exposed to moist heat (with reducing agent, thio or sulfhydryl group –SH), can be stretched--- fully extended b conformation. • After the reducing agent is removed, and an oxidizing agent is added to establish new disulfide bonds between pairs of cys. • The polypeptide chains revert to their a helical.---The new disulfide cross-linkages exert some torsion or twist on the bundles of a-helical coils in the hair fibers.
Structure of collagen • Like the a-keratins, collagen has evolved to provide strength, founded in connective tissue such as tendons, cartilage, the organic matrix of bone, the cornea of the eye • Collagen helix is left-handed and has 3 aa per turn. Gly-X –Pro or Gly-X-HyPro --- also form superhelical twisting in right-handed--- allow a very close packing to its three polypeptides---- tropocollagen molecule---provide tensile strength. tropocollagen
Structure of collagen fibrils • Collagen (Mr 300,000) is a rod-shaped molecule, about 3,000 Å long and only 15 Å thick. • Its three helically intertwined αchains may have different sequences, but each has about 1,000 amino acid residues. • Collagen fibrils are made up of collagen molecules aligned in a staggered fashion and crosslinked for strength. • alignment of the head groups of every fourth molecule produces striations 640 Å apart. • The tight wrapping of the chains in the collagen triple helix provides tensile (張力) strength greater than that of a steel wire of equal cross section.
Cross-kinked a chains of collagen molecules • The a chains of collagen and the collagen molecules of fibrils are cross-linked between Lys, HyLys or His --- dehydrohydroxylysinonorleucine. The increasing rigid and brittle character of connective tissue as people aged. • Mammal has more than 30 different collagen, in particular tissues: Osteogenesis imperfecta (成骨不全症--- abnormal bone formation in babies; Ehlers - Danlos syndrome (皮膚彈性過度症)--- loose joints (substitute Gly with larger R group, such as Cys or Ser)
Structure of silk fobroin • Fibroin, is produce by insects and spiders, mostly is b conformation --- rich in Ala and Gly. • Stabilized by H bonding between all pepetide linkages in each b sheet and by van der Waals interaction. • Silk does not stretch, because the conformation is already highly extended. However, the structure is flexible because the sheets are held together by numerous weak interactions rather than by covalent bonds such as the disulfide bonds in – a keratins.
Globular protein • Provides the structural diversity necessary for proteins. • Fording generates a compact peptide and provides the structural diversity --- wide array of biological functions, include enzymes, transport proteins, motor proteins, regulatory proteins, immunoglobulins, and proteins with many other functions.
Tertiary Structure of Small Globular Proteins: Myoglobin • The first breakthrough in understanding the three-dimensional structure of a globular protein came from x-ray diffraction studies by John Kendrew and his colleagues in the 1950s. • a relatively small (Mr 16,700); 153 AA. single iron protoporphyrin (heme). • It functions both to store oxygen and to facilitate oxygen diffusion in rapidly contracting muscle tissue. • 8 a helix interrupted by bend and all helices are right-handed. • Hydrophobic AA in the interior of protein--- compact (only 4 H2O)
Tertiary structure of sperm whale myoglobin (a) The polypeptide backbone, which highlights regions of secondary structure. The α-helical regions are evident. (b) A “mesh” image emphasizes the protein surface. (c) A surface contour image is useful for visualizing pockets in the protein where other molecules might bind.
Tertiary structure of sperm whale myoglobin (d) A ribbon representation, including side chains (blue) for the hydrophobic residues Leu, Ile, Val, and Phe. (e) A space-filling model with all amino acid side chains. Each atom is represented by a sphere encompassing its van deer Waals radius. The hydrophobic residues are again shown in blue; most are not visible, because they are buried in the interior of the protein.
The heme group • Present in myoglobin, hemoglobin, cytochromes, and many other heme proteins. • consists of a complex organic ring structure, protoporphyrin, to which is bound an iron atom in its ferrous (Fe 2+) state. • The iron atom has six coordination bonds, four in the plane and bonded to the flat porphyrin molecule and two perpendicular to His at position 93 and O2. • Heme in an oxygenated solution are rapidly oxidized from the ferrous (Fe2+ ), which is active in the reversible binding of O2, to the ferric (Fe3+ ) - does not bind O2.
Cytochrome c is a heme protein, covalently attach to protein, respiratory chain of mitochondria. Lysozyme is a hydrolytic enzyme (bacterial polysaccharides), abundant in egg white and human tears. Has 4 disulfide bonds. Ribonuclease, enzyme secreted by the pancreas into the small intestine, hydrolysis of certain ribonucleic acids. Has 4 disulfide bonds. Globular proteins have a variety of tertiary structures
Structural domains in the polypeptide troponin C • Supersecondary structures,also called motifsor simply folds, are particularly stable arrangements of several elements of secondary structure and the connections between them. • A single large motif may comprise the entire protein. • Domains: polypeptide with more than a few hundred AA often fold into two or more stable, globular units. • A domain from a large protein will retain its correct three-dimensional structure . • Structural domains in the troponin C (two calcium-binding domains).
Prominent folding rules of domains: • Hydrophobic interactions make a large contribution to the stability of protein structures. • a helices and b sheet generally are found in different structure layers • Adjacent peptide segment in the primary sequence are stacked to each other in the folded structure. • Connections between elements of secondary structure cannot cross or form knots. • The b conformation is most stable when the individual segments are twisted slightly in a right-handed sense.
Stable folding patterns in proteins (a) • Hydrophobic interactions make a large contribution to the stability of protein structures. Burial of hydrophobic amino acid R groups so as to exclude water requires at least two layers of secondary structure. • Two simple motifs, the b-a-b loopand the a-a corner, create two layers.