Retina
Retina. Transducing and Editing Patterns of Radiant Energy into Neural Signals. Retina: General. The retina is the innermost layer of the eye, located between the choroid and the vitreous


Retina
E N D
Presentation Transcript
Retina Transducing and Editing Patterns of Radiant Energy into Neural Signals
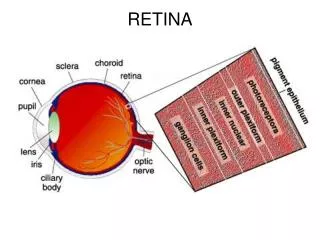
Retina: General • The retina is the innermost layer of the eye, located between the choroid and the vitreous • The retina extends from the optic nerve head to the ora serrata and is continuous with the layers of the ciliary epithelium, with which it shares embryologic origin • The retina is derived from neural ectoderm and consists of an outer pigmented layer, the retinal pigment epithelium, (RPE) derived from the outer layer of the optic cup, and the neural retina (all retinal layers other than the RPE) derived from the inner layer of the optic cup
Retina: General 2 • The pigmented RPE layer is tightly attached to the choroid throughout, but the neural retina is attached to the RPE only in a peripapillary ring around the optic nerve head and at the ora serrata • The retina is the site of the transformation of light energy into a neural signal and contains the first three cells (photoreceptor, bipolar, and ganglion cells) in the visual pathway • Other retinal cells—horizontal cells, amacrine cells, and interplexiform neurons—modify and integrate the signal before it leaves the eye
Retinal Histology • Examined with the light microscope, the retina has a laminar appearance with 10 “layers” (these include a layer of pigmented epithelium, 3 layers of neuronal cell bodies, between which lie their processes and synapses) • First, we will consider the epithelial layer and the types of function of the neural cells; the following section will consider the components of each of the 10 retinal layers
Outer nuclear layer Inner nuclear layer Ganglion cell layer
Retinal Pigment Epithelium • The retinal pigment epithelium (RPE), the outermost retinal layer, is a single cell thick and consists of pigmented hexagonal cells • These cells are columnar at the posterior pole and are even longer, narrower, and more densely pigmented in the macular area • The cells become larger and more cuboidal near the ora serrata • The basal aspect of the cell is adjacent to the choroid, and the apical surface faces the neural retina
Retinal Pigment Epithelium 2 • The basal aspect of the cell is adherent to its basement membrane, which is part of Bruch’s membrane, so its attachment to the choroid is strong • The RPE cells contain numerous melanosomes, pigment granules that extend from the apical region into the middle of the cell and obscure the nucleus • RPE pigment density varies across the retina and gives the fundus its mottled appearance • Melanin is densest in the RPE cells in the macula and at the equator
Retinal Pigment Epithelium 3 • Other pigmented bodies, lipofuscin granules, contain degradation products of phagocytosis and increase in number with age • The cell cytoplasm also contains rough and smooth endoplasmic reticulum, Golgi apparatus, mitochondria, and lysosomes • The apical portion of the RPE cell consists of microvilli that extend into the layer of photoreceptors, enveloping the specialized outer segment tips • No cellular junctions connect the RPE and photoreceptors – a potential space, the subretinalspace separates the RPE and the photoreceptor cells
Rod outer segment RPE villus RPE sheath Pigment granules in RPE apical portions
Retinal Pigment Epithelium 4 • Terminal bars, zonula occludens and zonula adherens, join the RPE cells near their apices • Desmosomes are present throughout the layer, and gap junctions allow for electrical coupling, providing a low-resistance pathway for the passage of ions and metabolites
Photoreceptor Cells • The photoreceptor cells, the rods and cones, are specialized sense cells containing photopigment that absorbs photons • The cells were originally named for their shapes, but the name does not always reflect the shape, particularly for the cones • A more significant distinction is the level of illumination over which each is active: rods are more active in dim illumination and cones in well-lit conditions
Photoreceptor Cells • Visual pigments are activated on excitation by light – 11-cis-retinal is isomerized to all-trans-retinal (vitamin A is all-trans-retinol) • A small amount of energy is released as a result of the isomerization and this begins the process of transduction (changing one form of energy into a different form)
RPE-Neural Retina Interface • Several factors help maintain the close proximity of the RPE and the photoreceptor outer segments • Passive forces, such as: • Intraocular pressure (IOP) • Osmotic pressure • Fluid transport across the RPE • Presence of the vitreous • Interdigitations between the RPE microvilli and the rod and cone outer segments • Material in the extracellular space between the two layers
RPE-Neural Retina Interface 2 • Recent studies have identified an organized honeycomb-like structure, the interphotoreceptor matrix (IPM) occupying the space between the RPE and the photoreceptors • The outer segments are completely surrounded by the IPM and extend through openings in the meshwork • The constituents of the IPM around the rods differs from that surrounding the cones; those surrounding the cones are described as a “matrix sheath”
RPE-Retinal Neural Interface 3 • The IPM material is tightly bound (through molecular bonds) to both the RPE and the photoreceptors and may exceed the strength of the RPE cells – with force, RPE cells rupture, leaving pigment remnants attached to the photoreceptors • The IPM provides a means of exchange of metabolites and for other interaction between the two layers • In addition, the IPM may be responsible for orienting the photoreceptor outer segments for optimal light capture—the Stiles-Crawford effect
Retinal Detachment • However, when a retinal detachment occurs, the separation generally lies between the RPE and the photoreceptors • Retinal detachment separates the retina from its blood supply, and unless the layers are repositioned quickly the photoreceptors will die • An argon laser is used to photocoagulate the edges of the detachment, which prevents the detached area from enlargening and helps reposition the photoreceptors
Rod and Cone Composition • The rods and cones are composed of several parts: • The outer segment, containing the visual pigment molecules • A connecting stalk, the cilium • The inner segment, containing the metabolic apparatus • The outer fiber (very short in cones) • The cell body • The inner fiber, which ends in a synaptic terminal
Synaptic terminals Rod and cone cell bodies Cell body External limiting membrane Switch Photoreceptor cells
Rod and Cone Composition 2 • The outer segment is composed of a stack of membranous discs (600 to 1000 per rod) and is enclosed by the plasmalemma of the cell • The discs are stacked on top of one another and separated by an extradisc space • Visual pigment molecules are located within the disc membranes – a biochemical change (an isomerization) is initiated within these molecules when they absorb a light photon • The tip of the outer segment is oriented toward the RPE, and the base is toward the incoming light
Rod and Cone Composition 3 • A connecting stalk, a cilium, extends from the innermost disc, joining the inner and outer segment and acting as a conduit between them • It is a modified cilium consisting of a series of nine pairs of tubules from which the central pair, usually present in motile cilia, are missing • The plasmalemma around the outer segments is continuous across the cilium with that of the inner segment
Rod and Cone Composition 4 • The inner segment contains cellular structures and can be divided into two parts: the ellipsoid and the myoid • The ellipsoid is nearer the outer segment and contains numerous mitachondria • The part closer to the cell body, the myoid, contains other cellular organelles such as the endoplasmic reticulum and Golgi apparatus; protein synthesis is concentrated in this area
Rod and Cone Composition 5 • The myoid in the human retina does not have contractile capabilities, though the photoreceptors’ outer and inner segments maintain an orientation toward the center of the exit pupil of the eye • The outer fiber extends from the inner segment to the cell body, the portion containing the nucleus • The inner fiber is an axonal process that runs inward from the cell body, ending in a specialized synaptic terminal that contains synaptic vesicles • The photoreceptors synapse with both bipolar and horizontal cells
Synaptic terminals Rod and cone cell bodies Cell body External limiting membrane Switch Photoreceptor cells
Rod and Cone Morphology • In the rods, the plasmlemma (outer wall) is separate from the discs except for a small region at the base, where interdigitations of the plasmalemma form discs • The rest of the discs are free floating – not attached to the surrounding plasmalemma or to each other • When radiolabeled amino acids were injected into the rod inner segments, where protein is synthesized, it moved into newly formed discs in the outer segment • The labeled discs moved from the base to the tip, and the label finally appeared in phagasomes of the RPE
Rod and Cone Morphology 2 • This rod outer segment renewal system involves active processes in both the RPE and the outer segment • Most of the shedding of rod discs occurs in the early morning • The rod inner fiber terminates in a rounded, pear-shaped spherule • The internal surface of the spherule is invaginated, forming a synaptic complex that contains bipolar dendrites and horizontal cell processes • Each rod spherule synapses with 1 to 4 bipolars, and rods release the neurotransmitter glutamate
Rod and Cone Morphology 3 • The outer segment of the cones is also enclosed by a plasmalemma, but in the cones, the plasma membrane is continuous with the membranes forming most of the discs (accordion folds) • In many cases the discs at the outer segment base are wider than those at the tip, giving the characteristic cone shape, though foveal cones have a shape similar to that of the rods • The cone outer segment is shorter than that of the rod, and may not reach the RPE layer, however, tubular processes protrude from the apex of the RPE cell to surround the cone outer segment
Rod and Cone Morphology 4 • One of three cone visual pigment molecules is contained within the cone disc membrane, and each molecule is activated by the absorption of light in a specific range of the light spectrum • The maximum absorptions for the three cones occur at about 420 nm (blue), 531 nm (green), or 588 nm (red) • New discs are formed at the base of the inner segment, but in the cones, radioactive-labeled amino acid is free to diffuse throughout the cone outer segment due to the interconnectedness of the discs • Cone discs are shed periodically, often at the end of the day, and are phagocytized by the RPE
Rod and Cone Morphology 5 • The shape of the inner segment also contributes to the cone shape – the cone ellipsoid area is wider and contains more mitochondria than the rod • The outer fiber (from the inner segment to the cell body) is short and stout and may even be absent in the cone, so the cone nucleus is closer to the external limiting membrane than are the rod nuclei • The inner fiber terminates in a broad, flattened structure called a pedicle, which has several invaginated areas
Rod and Cone Morphology 6 • Cone pedicles make several types of synapses – triads are found within the invaginations, where central contact is made with the dendrite of a midget bipolar cell, flanked by contacts with the processes of two different horizontal cells • Also, contact with flat bipolar cells are made on the flat surface of the pedicle, near the synaptic invaginations • Gap junctions are located on the lateral expansions (telodendria or interphotoreceptor connections) of the pedicle and permit electrical communication between two cones, or between a cone and a rod • Like rods, cones use glutamate as a transmitter
Bipolar Cells • The bipolar cell is the second-order neuron in the visual pathway • Its dendrite synapses with photoreceptor and horizontal cells, and its axon synapses with ganglion and amacrine cells • Glutamate is its transmitter • Bipolar cells relay information from the photoreceptors to horizontal, amacrine and ganglion cells, and receive extensive synaptic feedback from amacrine cells
Bipolar Cells 2 • Eleven types of bipolar cells have been identified based on morphology, physiology, and dendritic contacts with the photoreceptors • All types of bipolars are associated with cones except for the rod bipolar – bipolars either connect to rods or cones, but never to both • The rod bipolar has a large body and several spiky dendrites – these are the only bipolars that contact rodsand they form the central element of a triad
Midget bipolars Diffuse small-field bipolars
Bipolar Cells 3 • The dendrites of a single rod bipolar contact 15 to 20 rods in central retina and up to 80 rods in the periphery • A triad consists of processes from two horizontal cells flanking a bipolar dendrite within an invagination in a photoreceptor; there is usually one triad per rod • The rod axon is large and unbranched and may not synapse directly with the ganglion cell but instead may synapse with amacrine processes, which then contact ganglion cells
Bipolar Cells 4 • Midget bipolars have a relatively small cell body and come in two varieties: 1) flat, and 2) invaginating • The flat midget bipolar ends in a flat expansion and makes contact only with the flat area of the cone pedicle – in the central retina, each flat midget bipolar contacts only one cone, and in the peripheral retina, each flat midget bipolar contacts two or three neighboring cones • A single cone pedicle may have as many as 500 contacts on its flat pedicle surface
Bipolar Cells 5 • The invaginating midget bipolar’s dendritic process is the central element in a triad and is flanked by two horizontal cell processes (from different horizontal cells) – this triad is located within an invagination of the cone pedicle • In the central retina, like the flat midget bipolar, one invaginating midget bipolar may contact only one cone, but in the periphery the bipolar may contact several cone pedicles • The axon of the invaginating midget bipolar cell synapses with the dendrite of a single midget ganglion cell and with amacrine processes
Bipolar Cells 6 • There are two types of diffuse cone bipolars and in the central retina they contact 5 neighboring cones and in the periphery about 10 to 15 neighboring cones • The blue cone bipolar synapses with three widely spaced blue cones (cones with their maximum absorption in the blue range of the spectrum) • The giant cone bipolars have a large dendritic tree and are divided into diffuse and bistratified based on the spatial distribution of their axon terminals
Ganglion Cells • The next cell in the visual pathway, the third-order neuron, is the ganglion cell – they can be bipolar (single axon and single dendrite) or multipolar (single axon and more than one dendrite) • Ganglion cell size varies greatly and they have been classified on the basis of cell body size, branching characteristics, termination of dendrites, and diameter of fiber and dendritic tree • Another, more functional, designation classifies ganglion cells based on the lateral geniculate body (LGN) layer in which they terminate
Ganglion Cells 2 • P cells terminate in the parvocellular layers (the four dorsal layers of the LGN) • The P1 ganglion cell, also called the midget ganglion cell, is the most common p cell – certain P1 midget cells are connected to only one midget bipolar cell, invaginating or flat, which in turn might be linked to a single cone photoreceptor – this provides a channel that processes high-contrast detail and color resolution • This arrangement is most likely to occur in the fovea
Ganglion Cells 3 • The P2 ganglion cell also terminates in the parvocellular layers, but has a densely branched compact dendritic tree that spreads horizontally • The M-type ganglion cell projects to the magnocellular layers (two ventral layers) of the LGN • The M cell has coarse dendrites and the dendritic tree enlarges from the central to peripheral retina • Each ganglion cell has a single axon, which emerges from the cell body and turns to run parallel to the inner surface of the retina (forming the nerve fiber layer); the axon uses glutamate as its neurotransmitter
Ganglion Cells 4 • The axons (1-2 million in each eye) come together at the optic nerve head and leave the eye as the optic nerve • The termination for about 90% of these axons is the LGN; others project to subthalamic areas involved in pupillary reflexes and circadian rhythm • The photoreceptors, bipolar cells, and ganglion cells constitute the direct visual pathway in the retina – other cells modify the signal carried by means of intraretinal cross-connections, provide feedback, or integrate retinal function – these modifying cells are the horizontal, amacrine and interplexiform cells • In general, the retinal neural network signals change in the retinal image and ignores static features
Horizontal Cells • The horizontal cell transfers information in a horizontal direction parallel to the retinal surface • It has one long process, or axon, which terminates in the innerplexiform layer, and several short dendrites, which terminate in the outer plexiform layer • Horizontal cells synapse with photoreceptors, bipolar cells, and other horizontal cells – horizontal cells are thought to contact bipolar cells lying some distance from the photoreceptor that directly activates the horizontal cell