Download

1 / 45
450 likes | 700 Views
I count 1, 2, 3, 4 , 5 p53 mutants and…oh…~300,000 wild-type DNA molecules. DEVELOPING TUMOR-ASSOCIATED SOMATIC MUTATIONS AS BIOMARKERS OF CANCER RISK. Barbara Parsons Division of Genetic and Reproductive Toxicology National Center for Toxicological Research. ACB-PCR.

E N D
I count 1, 2, 3, 4 , 5 p53 mutants and…oh…~300,000 wild-type DNA molecules. DEVELOPING TUMOR-ASSOCIATED SOMATIC MUTATIONS AS BIOMARKERS OF CANCER RISK Barbara Parsons Division of Genetic and Reproductive Toxicology National Center for Toxicological Research ACB-PCR
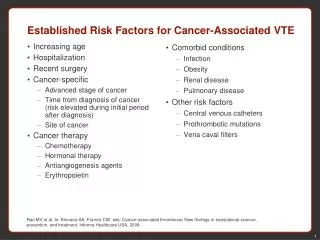
Regulatory Research Need • Albertini et al. (2003) The use of non-tumor data in cancer risk assessment: reflections on butadiene, vinyl chloride and benzene. Regulatory Toxicology and Pharmacology 37: 105-132 “Reliance on animal tumor incidence data with its inherent need to extrapolate to humans introduces considerable uncertainty into the risk assessment process.” • To determine whether/how quantitative measurement of tumor-associated mutation can be used to improve cancer risk assessment (first steps toward validating the use of tumor-associated somatic mutation as a biomarker for cancer risk). Research Goal
Advantages of DNA-based measurement of tumor-associated mutations • EPA’s Cancer risk assessment guidelines indicate risk assessment should be based on an understanding of chemical mode-of-action and key events in carcinogenesis. • Somatic mutations are considered the earliest biomarkers of chemical effect (tumor-associated mutations considered early biomarkers of disease state). • A large literature supports the interpretation that mutation in oncogenes and tumor suppressor genes are key events in carcinogenesis. • Tumor-associated mutations considered more relevant biomarkers of chemical effect than mutations in surrogate reporter genes (hprt, lacI, and Tk).
Tumor-associated mutations are acceptedas key events in carcinogenesis • “Colorectal cancers arise through a gradual series of histological changes - the adenoma-carcinoma sequence-resulting from specific genetic “hits” at a handful of oncogene and tumour suppressor genes.” Fodde et al., Nature Reviews Cancer 1, 55-67 (2001). • Hallmarks of Cancer by Hanahan and Weinberg, Cell 100, 57-70 (2000) indicates several acquired characteristics of tumors can be caused by specific point mutations – Ras and p53. • Data from transgenic p53 and ras rodents accepted in carcinogenicity testing. • At the 2006 EMS meeting, Dr. Channa Keshava indicated EPA weighs oncogene/tumors suppressor gene mutation more heavily than reporter gene mutational data in a weight-of-evidence approach.
Allele-specific Competitive Blocker-PCR (ACB-PCR) * Priming strategy for K-RAS codon 12 GGT GTT mutation
Quantifying Mutant Fraction Synthesize first-round PCR product (Pfu polymerase) Aliquot, quantify all unknowns and standards Isolate genomic DNA from tissue ACB-PCR of duplicate MF standards, no mutant, no DNA controls, and ~24 unknowns in parallel, per experiment Polyacrylamide gel electrophoresis, measure fluorescence, use MF standards to a construct a standard curve, calculate the MF (ratio of mutant to wild-type sequence of unknowns, repeat assay 3 times R2=0.9924 no mutant control
ACB-PCR assays developed to date • Mouse H-Ras codon 61 CAA to AAA mutation • Mouse p53 codon 270 CGT to TGT mutation • Mouse K-RAS codon 12 GGT to TGT mutation • Human K-RAS codon 12 GGT to GTT mutation • Human K-RAS codon 12 GGT to GAT mutation • Rat K-Ras codon 12 GGT to GTT mutation • Rat K-Ras codon 12 GGT to GAT mutation • The ACB-PCR methodology has been developed for seven different mutations, each with a sensitivity of 10-5 (i.e., detects 3 mutant DNA molecules among 300,000 wild-type molecules). • Limit of detection 10-5, although the value calculated from the standard curve (values below 10-5) is usually used in statistical analyses.
Principles for using non-tumor data in cancer risk assessment Albertini et al., 2003 ACB-PCR was used to investigate relationship between DNA adducts, H-Ras codon 61 CAA to AAA mutation, and tumor induction in newborn B6C3F1 and C57BL/6 mice treated with 0.3 mol 4-aminobiphenyl “The more we understand about the applied dose metabolism mutation effect continuum, the more confidence we can have in a cancer risk assessment” • Tumor data was available from a newborn mouse assay, where tumor induction by 0.3 umol of 4-ABP was scored at 1 year . • B6C3F1 and C57Bl/6 mice were given i.p. injections of 4-ABP (or DMSO vehicle) on postnatal days 8 and 15 days (12 mice per treatment group). • Some animals sacrificed 24 hours after the second dose for liver DNA adduct analyses (Dr. Fred Beland). • Livers were harvested 8 months after treatment - normal by gross pathological examination. • DNA isolated from livers, made 1st round PCR product, then analyzed for H-Ras codon 61 CAA to AAA mutation by ACB-PCR.
Induction of mutation and correlation with strain susceptibility P=0.0515 P=0.0008 Molecular Carcinogenesis 42:193-201 (2005)
Can liver H-Ras MF at 8 monthspredict tumor development at 1 year? Spontaneous H-Ras codon 12 CAA to AAA MF in mouse liver is closer to 10-5 than 10-7 or 10-8 as was expected based on reporter gene MFs.
Principles for using non-tumor data in cancer risk assessment Albertini et al., 2003 Compared the levels of two different K-Ras codon 12 mutations in livers of Big Blue rats treated with N-hydroxy-2-acetylaminofluorene in an attempt to distinguish between chemical-induced induction of de novo mutation vs. chemically-induced amplification of pre-existing mutation.G T mutation, the predominant mutagenic specificity of N-OH-AAFG A mutation, the most common spontaneous mutation • “Non-tumor data is critical for determining mechanism/mode of action” • Tao Chen et al. characterized lacI mutation in six week old Big Blue rats given four i.p. injections of 25 mg/kg N-OH-AAF or the vehicle control at four day intervals. Environmental and Molecular Mutagenesis (2001) 37: 203-214. • Page McKinzie measured the induction of K-Ras codon 12 GGT GTT and GGT GAT mutation in liver DNA using ACB-PCR.
Comparison of lacI and K-Ras MF measurementssuggests N-OH-AAF may cause amplification of pre-existing mutation in rat liver lacI K-Ras McKinzie et al. (2006) Mutagenesis 21: 391-397
Principles for using non-tumor data in cancer risk assessment Albertini et al., 2003 • “Non-tumor data can improve the validity of extrapolating from high doses to low doses” • Sunlight preferentially induces CT mutation at PyrmeCpG target sites, codon 270 is such a target. • This mutation has a dominant negative phenotype. • The codon 270 CGT to TGT mutation has been found in a significant percentage (20-60%) of mouse SCCs induced by UV-B or SSL. • p53 mutation is viewed as a key causal event in the development of sunlight/UV-B induced squamous cell carcinomas (SCCs) in both mouse and human. Conducted a dose-response assessment, measuring the induction of p53 codon 270 CGT to TGT mutation by simulated solar light (SSL).
National Toxicology Program Center for Phototoxicology Filtered 6.5 kW xenon arc light mimics terrestrial exposure to sunlight • SKH-1 hairless mice receive sub-toxic doses of 0, 6.85, 13.70, or 20.55 mJ∙CIE/cm2 per day, five days per week (equivalent to 0, 7.5, 15 or 30 minutes of exposure to terrestrial sunlight) • p53 mutation measured in skin after 4, 16, or 28 weeks of exposure.
Measured the p53 codon 270 CGT to TGT mutational response induced by 4 different doses of SSL – after three different durations of exposure
Dose-response analysis and measurement of spontaneous p53 MF in skin Trend, p=0.282 Trend, p<0.001 Trend, p<0.001 P = 0.008
The increase in p53 MF with cumulative dose is highly-significant P = <0.0001 MF = 10α100.0016*d, where α is the spontaneous MF at a particulartimepoint and d is the dose of SSL in mJ CIE/cm2
MF=0.5 p53 MF measurements in skin tumors All skin tumors have significant levels of p53 codon 270 mutation, but the mutation appears to represent a sub-population of tumor cells.
Comparison of p53 MF in skin and skin tumors Tumors induced by 32 weeks of exposure at the high dose had significantly more p53 mutation than skin exposed for 28 weeks at the high dose (t-test, P = 0.0006).
Future research efforts on p53 project • Isolating DNA from replicate needle biopsies from 4 and 7 month animals (medium and high doses). • Trying to use ACB-PCR to characterized the frequency distribution of p53 mutation within skin. • Effort to describe changes in p53 mutant clone size during tumor development. • Hypothesize that p53 mutation is being removed at 7 months but some clones, those on the path to tumor development, must be getting larger
Principles for using non-tumor data in cancer risk assessment Albertini et al., 2003 • “Endogenous levels of biomarkers can provide information for risk assessment” • “Biomarkers must be selected and interpreted in light of what is known about mechanism of action” • K-RAS mutation is a “key event” in the development of sporadic colon cancer • K-RAS mutation has been detected in normal colon and in all stages of colon cancer development • K-RAS mutation causes constitutive mitogenic signaling. • K-RAS mutation induces expression of VEGF which induces angiogenesis. • Detection of K-RAS codon 12 GGT to GTT mutation in sporadic colonic adeno- carcinoma has been associated with poor prognosis. • Quantified the levels of K-RAS codon 12 GAT and GTT mutation in the colonic mucosa of human and rat. Determined the levels of K-RAS mutation in human tissues corresponding to pathological stages of colon tumor development, information that may be used to develop mathematical models of carcinogenesis based on oncogene/tumor suppressor gene MFs.
Literature review of APC and K-RAS mutation prevalence in sporadic colon cancer GAT mutation in ~20% of colonic adenocarcinoma GTT mutation in ~7% of colonic adenocarcinoma Fodde et al. (2001) Nature Reviews Cancer 1:55-67
Mutation theory and modeling sporadic colon tumor development Vogelgram Luebeck and Moolgavkar (2002) PNAS Assumes clonal amplification after acquiring multiple mutations
ACB-PCR analysis ofhuman colon tissues • Collected fresh tissue from the Central Arkansas Veterans Healthcare System (CAVHS) hospital, as well as fresh-frozen tissues from the Cooperative Human Tissue Network (CHTN). • 77 samples analyzed in triplicate for K-RAS codon 12 GAT and GTT mutation (462 data points). • Samples analyzed to date: Normal (individuals w/o colon ca.) 2 Tumor distal mucosa (5 or more cm from tumor) 17 Tumor proximal mucosa (2 to 5 cm from tumor) 14 Tumor margin (normal tumor-adjacent) 10 Adenoma 12 Adenocarcinoma 21 • Additional samples, including ACF remain to be examined.
GAT GTT Adenocarcinoma Tumor distal mucosa Tumor proximalmucosa Tumor margin Normal Adenoma
Significant Findings • Normal-appearing colon contains a significant level of K-RAS mutation (with GAT>GTT). • Significant differences in GAT and GTT MF were found in the different tissues (Kruskal-Wallis ANOVA on ranks, P=0.013 and P=0.006) with GAT MF in adenoma and adenocarcinomas significantly higher than in mucosa and GTT MF in adenoma significantly higher than in mucosa and adenocarcinoma. • Many tumors carry both K-RAS mutations as sub-populations.
Significant Findings • The decrease in K-RAS GTT MF during tumor progression suggests that tumors are composed of more than one clone of tumor cells (K-RAS mutant and non-K-RAS mutant) and that during progression to adenocarcinoma, K-RAS mutant cells are either dying or other clone(s) are proliferating faster.
It is widely accepted that tumors are monoclonal in origin (arise from a single initiated cell). This conclusion is based on studies from the 1970s which showed the same X-chromosome-linked marker was inactivation in all cells of a tumor (colon tumors). Amazon.com NCI Teacher’s Guide, “Understanding Cancer”…cancer cells are indigenous cells—abnormal cells that arise from the body's normal tissues. Furthermore, virtually all malignant tumors are monoclonal in origin, that is, derived from a single ancestral cell that somehow underwent conversion from a normal to a cancerous state. One Renegade Cell (Science Masters) (Paperback) by Robert A. Weinberg (Author)
Evidence supporting polyclonal tumor origin • 76% of adenomas in an XO/XY FAP chimeric woman with average patch size of 1.48 crypts were polyclonal. [Novelli et al., Science 272:1187-1190 (1996)]. • Novelli demonstrated that X-inactivation happens early enough in develoment that the human colon has an average patch size of 450 crypts, meaning there is a strong bias for detecting monoclonality. [Novelli et al., PNAS 100:3311-3314 (2003)]. • Min+/- heterozygous, ROSA26 chimeric mice also shows high frequency of polyclonal adenomas. [Thliveris et al., PNAS 102:6960 (2005)]. • In mouse, active short-range interaction between multiple initiated crypts is responsible for polyclonality of adenomas.Mutant crypts may cause elevated proliferation rate of stem cells in adjacent crypts or interaction between initiated clones may favor tumor survival and development of polyclonal adenomas. [Leedham et al., Cell Proliferation 38: 387 (2005), Thliveris et al., PNAS 102:6960 (2005)]. • This issue is currently being re-examined. A recent survey of the literature (since 1997) found 13 reports that conclude tumors are monoclonal and >20 reports with evidence of polyclonality for 10 different tumor types.
Interpreting Evidence of Tumor Clonality Monoclonal origin Polyclonal origin Thliveris et al., PNAS 102:6960 (2005). • Cannot prove a negative, so reports of monoclonality should be interpreted cautiously. Reports of polyclonality, perhaps, should be given more weight. • Conceptually, the best evidence of polyclonality will be obtained using markers heterogeneously distributed throughout the normal tissue and present as sub-populations within tumors.
Principles for using non-tumor data in cancer risk assessment Albertini et al., 2003 Comparing spontaneous levels of K-Ras mutation in rat and human colonic mucosa • “Endogenous levels of biomarkers can provide information for risk assessment” *Mean age 67 years #38 week-old rats (Page McKinzie) Colon of a 38 week-old rat has a K-RAS MF 7 to 15 times the of a 67 year-old man
Can we model carcinogenesis based on the frequency and spatial distribution of different tumor-associated mutations? K-RasGTT K-RasGAT APC Frequency and distribution of mutation relates to frequency of adenoma formation Adenoma Adenocarcinoma Michael A. Newton, University of Wisconsin Seminar, On Polyclonality of Intestinal Tumors describes the “Random Collision Hypothesis” originally proposed by Armitage http://sprott.physics.wisc.edu/Chaos-Complexity/newton06.pdf
Summary • Developing oncogene and tumor suppressor gene mutations as biomarkers of cancer risk. • Demonstrated that spontaneous levels of K-Ras and p53 mutation are much higher than expected based on comparisons with neutral reporter gene measurements. • Demonstrated statistically-significant induction of oncogene and tumor suppressor gene mutation using model mutagens (4-ABP, N-OH-AAF, SSL). • ACB-PCR measurements will be useful for establishing chemical mode-of-action (distinguishing de novo mutation from amplification of pre-existing mutation). • Used ACB-PCR to describe the dose response relationship between SSL exposure and induction of p53 codon 270 CGT to TGT MF in mouse skin. • Somatic oncogene and tumor suppressor gene mutations are frequently represented as sub-populations within tumors. • Analysis of K-RAS MF in human and rat colon illustrates how ACB-PCR can be used to inform species extrapolation (better understanding of how the rodent model compares to human). • Quantitative analysis of tumor-associated mutation through the different stages of carcinogenesis can provide a scientific framework for the use of tumor-associated mutation as a metric of cancer risk. • In the future, these measurements may be applied on a case-by-case basis, as part of the weight of evidence approach to cancer risk assessment.
Studies Planned or in Progress • Pilot dose-response study in collaboration with Jeff Ross (NHEERL/EPA) to measure K-Ras TGT mutation in lung tissue of A/J mice exposed to a range of doses of benzo[a]pyrene (low to tumorigenic doses). Investigating the utility of ACB-PCR in measuring a response at low dose with a short expression time and the relationship between DNA adducts and mutation. • Planning a collaboration with Harvey Clewell (CIIT Centers for Health Research) to measure dose-dependent induction of a p53 mutation in the nasal mucosa of formaldehyde–treated rats. CIIT will collect data on the dose-dependent induction of toxicity-induced cell proliferation. We will learn whether the dose-dependent appearance of p53 mutation coincides with that of formaldehyde-induced cytotoxity and compensatory cell proliferation.
This research was supported by: NCTR NTP Center for Phototoxicology Rat K-Ras Studies Page McKinzie Robert Delongchamp John Latendresse Tao Chen Human K-RAS Study Kathy Marchant Robert Delongchamp Page McKinzie Tucker Patterson Ralph Broadwater Laura Lamps Lawrence Kim p53 SSL Study Tracie Verkler Robert Delongchamp Alan Warbritton Letha Couch Barbara Miller Peggy Webb Paul Mellick Paul Howard Martha Moore Director, Division of Genetic and Reproductive Toxicology
References Ablertini, R., H. Clewel, M.W. Himmelstein, E. Morinello, S. Olin, J. Preston, L. Scarano, M.T. Smith, J. Swenberg, R. Tice, and C. Travis (2003) The use of non-tumor data in cancer risk assessment: reflections on butadiene, vinyl chloride, and benzene. Regulatory Toxicology and Pharmacology 37: 105-132. Chen, T., R.A. Mittelstaedt, S.D. Shelton, S.B. Dass, M.G. Manjanatha, D.A. Casciano, and R.H. Heflich (2001) Gene- and tissue-specificity of mutation in Big Blue rats treated with the hepatocarcinogen N-hydroxy-2-acetylaminofluorene. Environmental and Molecular Mutagenesis 37: 203-214. Fearon, E.R. and B. Vogelstein (1990) A genetic model for colorectal tumorigenesis. Cell 61: 759-767. Fodde, R., R. Smits, and H. Clevers (2001) APC, signal transduction and genetic instability in colorectal cancer. Nature Reviews Cancer 1: 55-67. Hanahan, D., and R.A. Weinberg (2000) The hallmarks of cancer. Cell 100: 57-70. Leedham, S.J., S. Schier, T. Thliveris, R.B. Halberg, M.A. Newton, and N.A. Wright (2005) From gene mutations to tumours – stem cells in gastrointestinal carcinogenesis. Cell Proliferation 38: 387-405. Luebeck, E.G., and S.H. Moolgavkar (2002) Multistage carcinogenesis and the incidence of colorectal cancer. PNAS 99: 15095-15100.
References McKinzie, P.B., R.R. Delongchamp, T. Chen, and B.L. Parsons (2006) ACB-PCR Measurement of K-ras codon 12 mutant fractions in livers of Big Blue rats treated with N-hydroxy-2-acetylaminofluorene. Mutagenesis 21: 391-397. McKinzie, P.B., R. Delongchamp, R.H. Heflich, and B.L. Parsons (2001) Prospects for applying genotypic selection of somatic oncomutation to chemical risk assessment. Mutation Research 489: 47-78. McKinzie, P.B., and B.L. Parsons (2002) Detection of rare K-ras codon 12 mutations using allele-specific competitive blocker PCR. Mutation Research 517: 209-220. Newton, M.A (2006) On polyclonality of intestinal tumors. University of Wisconsin. Chaos and Complex Systems April 2006. http://sprott.physics.wisc.edu/Chaos-Complexity/newton06.pdf Novelli, M.R., J.A. Williamson, I.P.M. Tomlinson, G. Elia, S.V. Hodgson, I.C. Talbot, W.F. Bodmer, N.A. Wright (1996) Polyclonal origin of colonic adenomas in an XO/XY patient with FAP. Science 272: 1187-1190. Novelli, M., A. Cossu, D. Oukrif, A. Auaglia, S. Lakhani, R. Poulsom, P. Sasieni, P. Carta, M. Contini, A. Pasca, G. Palmieri, W. Bodmer, F. Tanda, and N. Wright (2003) X-inactivation patch size in human female tissue confounds the assessment of tumor clonality. PNAS 100: 3311-3314.
References Parsons, B.L., F.A. Beland, L.S. Von Tungeln, R.R. Delongchamp, P. Fu, and R.H. Heflich (2005) Levels of 4-aminobiphenyl-induced somatic H-ras mutation in mouse liver DNA correlate with potential for liver tumor development. Molecular Carcinogenesis 42:193-201. Parsons, B.L., R.R. Delongchamp, F.A. Beland, R.H. Heflich (2006) Levels of H-ras codon 61 CAAAAA mutation: response to 4-ABP-treatment and Pms2-deficiency. Mutagenesis 21: 29-34. Parsons, B.L. and R.H. Heflich (1997) Genotypic selection methods for the direct analysis of point mutations. Mutation Research, 387: 97-121. Parsons, B.L. and R.H. Heflich (1998) Detection of a mouse H-ras codon 61 mutation using a modified allele-specific competitive blocker PCR genotypic selection method. Mutagenesis 13: 581-588. Parsons, B.L. P.B. McKinzie, and R.H. Heflich (2005) ACB-PCR detection of rare point mutation. In: Molecular Toxicology Protocols (Methods in Molecular Biology Series), P. Keohavong and S. Grant (eds.), Humana Press, Totawa, NJ.
References Thilveris, A.T., R.B. Halberg, L. Clipson, W.F. Dove, R. Sullivan, M.K. Washington, S. Standhope, and M.A. Newton (2005) Polyclonality of familial murine adenomas: analyses of mouse chimeras with low tumor multiplicity suggest short-range interactions. PNAS 102: 3960-6965. Verkler, T.L., L.H. Couch, P.C. Howard, and B.L. Parsons (2005) Quantifying levels of p53 mutation in mouse skin tumors. Environmental and Molecular Mutagenesis 45: 427-43.
Next steps:Determine mutant clone size • Perform ACB-PCR “fluctuation test” on mucosa to determine the frequency and size distributions of mutant clones (multiple small biopsies). • Convert this information to number of K-RAS GAT and GTT mutant and wild-type crypts. • Approximate the frequency of APC mutant clones from their abundance relative to K-RAS ACF in the literature (for FAP and sporadic). • Mathematically (?) relate the expected co-occurrence of mutant crypts with the probability of developing K-RAS mutant sporadic colon cancer and determine whether this relationship predicts the increased probability of cancer in FAP patients when the increased number of APC crypts is incorporated.
Potential implications of polyclonal tumor formation • Mutant clones may contribute transiently to tumor development. • Cannot interpret the most abundant mutation in a tumor as the “initiating” mutation. • Possibility that a few specific mutations are universally involved in the development of a particular tumor type. • Can’t correctly model tumor development without an accurate understanding of the process! • Can’t accurately interpret changes in the frequency of tumor-associated mutations unless their role on the process of carcinogenesis is understood. • Quantitative measurements of tumor-associated mutation provide new opportunities to investigate the earliest events in carcinogenesis.