Download

1 / 37
370 likes | 660 Views
Metanogeenid, Archaeoglobus ja Ferroglobus Kuuluvad arhede hõimkonda Euryarchaeota. Kuuluvad hõimkonda Euryarchaeota. Archaeoglobusel on nii metanogeenide kui ka krenarhede jooni. Metanogeenid. Suur ja kirju rühm, mida ühendab 3 joont: 1. metaani moodustamine, 2. range anaeroobsus,

E N D
Metanogeenid, Archaeoglobus ja Ferroglobus Kuuluvad arhede hõimkonda Euryarchaeota
Kuuluvad hõimkonda Euryarchaeota. Archaeoglobusel on nii metanogeenide kui ka krenarhede jooni.
Metanogeenid Suur ja kirju rühm, mida ühendab 3 joont: 1. metaani moodustamine, 2. range anaeroobsus, 3. kuuluvus arhede hulka.
Levik looduses ja füsioloogia. Okupeerivad anaeroobseid paiku looduses:muda, seedekulgla, sood, üleujutatud muld, metaantankid, mäletsejate loomade vats, termiitide sool, igemetaskud.Elavad sageli koos teiste arhedega. Metanogeenid on väga hapnikutundlikud. Näiteks Methanosarcina hukub laboris puhaskultuuris hapniku juuresolekul 4 minuti jooksul. Looduses nad nii tundlikud ei ole, sest teised bakterid kaitsevad neid, tarvitades hapniku ära. Metanogeene on seetöttu näiteks isoleeritud hambaaukudest ja hambakatust.
Anaeroobsetes tsoonides pidurdab metanogeneesi nitraatide, sulfaatide ja Fe3+ esinemine, sest anaeroobsed hingajad törjuvad siis metanogeenid välja. Näiteks sulfaatsete hingajate afiinsus H2 suhtes on palju suurem, kui metanogeenidel. Setõttu “saavad” sulfaatsed hingajad vesiniku endale. Ka inimese soolestikus saab sulfaadirikka toiduga viia alla metaani teket: H2 oksüdeeritakse siis H2S tekkega ja metaani moodustub vähem.
Mitmetes looduspaikades ei söltu metanogeenid kääritajate mikroobide poolt toodetud vesinikust ja atsetaadist. Nii näiteks on termofiilseid metanogeene isoleeritud piirkondadest, kus H2 on vulkaanilise päritoluga. NB! Hiljuti on avastatud, et metanogeenid saavad metaani tekkel oksüdeerida ka rauda. Seega võivad nad osaleda anaeroobses raudkonstruktsioonide biokorrosioonis.
Levik looduses ja füsioloogia. Atmosfääris on metaani [c] 1.7 ppm ja ta on C sisaldavatest gaasidest atmosfääris CO2 järel teisel kohal. Metaani hulk atmosfääris suureneb pidevalt, ca 1% aastas ja metaan on ka üks kasvuhooneefekti pöhjustaja, kuna neelab infrapunast kiirgust. Arvatakse, et ca 74% sellest tekkivast metaanist on mikrobioloogilist päritolu.
Enamus metanogeene elab nn tavalistes tingimustes: tavalisel temperatuuril ega vaja soola või kõrget keskkonna happesust. Erandiks on Methanopyrus, kes on termofiil. Methanopyruson evolutsiooniliselt vana perekond. Tema uurimine on näidanud, et tal on tsütoplasmas väga kõrges kontsentratsioonis tsüklilist 2,3-difosfoglütseraati, mis võiks olla termostabilisaator valkudele ja DNA-le. Ka on tal leitud eeterlipiidid, mille süsivesinikahel on küllastumata (teistel arhedel küllastatatud). Methanopyrus isoleeriti “mustast suitsetajast” 2000 m sügavuselt ookeani põhjast. Varem arvati, et sellistes tingimustes (kuumus) ei saa metaani moodustuda.
Metanogeneesi substraadid Metanogeensete bakterite puhaskultuuridega on näidatud, et vähemalt 10 erinevat substraati on metaaniks konverteeritavad. Need substraadid saab jagada 3 rühma: 1.CO2-tüüpi substraadid. Siia kuuluvad CO2 formiaat, ja CO. Elektronid selle redutseerimiseks tulevad vesinikult, püruvaadilt või teatud alkoholidelt; 2.Metüülühendid. Metanool, metüülamiin, dimetüülamiin, trimetüülamiin, metüülmerkaptaan, dimetüülsulfiid. 3.Atsetaat. Vesinik on kõide levinuim oksüdeeritav substraat metanogeenidele.
Metaani moodustumisel CO2-st redutseeritakse ta metaanini ja tavaliselt saadakse elektronid selleks H2 oksüdeerimisest. CO2 + 4H2 CH4 + 2H2O
Metaani teke vesinikust ja CO2-st Metanogeenid sisaldavad mitmeid unikaalseid koensüüme, mis on seotud metaani biosünteesi biokeemiaga. Metanofuraan, metanopteriin ja CoM on C-aatomi kandjad selle redutseerimisel CO2-st metaanini. Elektronid CO2 redutseerimiseks tulevad H2 oksüdatsioonist ja mönede teiste rühmade oksüdatsioonist. Neid elektrone kannavad CoF420 ja 7-merkaptoheptanoyltreoniinfosfaat (HTP).
Metanofuraan (MFR) on formüüljäägi kandja metanogeneesil. See on vajalik CO2 algseks sidumiseks ja redutseerimiseks formüülmetanofuraaniks. R-külgahel on küllalt keerulise struktuuriga
Tetrahüdrometanopteriin (H4MPT) on 1C-kandja. Ta kannab C1-jääki selle redutseerumisel formüülrühmast (-CHO) kuni metüülrühmani (-CH3). Ta struktuur on lähedane foolhappe struktuurile ja funktsioonile. R-külgahelat ei ole välja joonistatud
CoMon 2-merkaptoetaansulfoonhape. Osaleb metaani teke lõpureaktsioonis. CoM toimib terminaalse C-kandjana metanogeneesis. Metüül-CoM (paremal) redutseeritakse metaaniks.
Co F420 on koensüüm, mis annab metanogeenide rakkudele autofluorestsentsi. Teda on metanogeenides rohkesti ja teistes organismides ka, kuid palju vähem. F420 osaleb 2-elektronilistes ülekannetes ja tal on NAD-ga sarnane funktsioon.
F420 on elektroni doonor metanogeneesil. Ta on seotud hüdrogenaasiga, formiaadi dh-ga, NADH reduktaasiga, CO dh-ga jmt ensüümidega. Võtab vastu nende ensüümide poolt substraatidelt ärastatud elekronid ja suunab need metaani tekke ratta. Huvitaval kombel on see koensüüm olemas ka sulfaate redutseerival arhel Archaeoglobus.
Metanogeenid sisaldavad ka ferredoksiini ja cytb ja cytc. Tsütokroomid funktsioneerivad elektroni kandjatena metanogeneesil, mis toimub metanooli, metüülamiini ja atsetaadi baasil. Nad osalevad nende molekulide oksüdatsioonis. Cyt-d puuduvad näiteks Metanosphaera’l, kes kasutab metanogeneesilainult H2 + CO2.
Metüülrühma sisaldavatel ühendite puhul on metaani tekkeks kaks võimalust. Esiteks, metüülrühm redutseeritakse metaanini, kasutades eksogeense redutseerijana näiteks H2. CH3OH + H2 CH4 + H2O Kui aga H2 pole, siis oksüdeeritakse osa substraadi molekule CO2-ks ja ärastatud elektrone kasutatakse teise osa substraadi molekulide metüülrühma redutseerimiseks metaanini. 4CH3OH 3CH4 + CO2 + 2H2O Looduses moodustub metüleeritud ühendeid rohkesti. Mere- ja järvemudas vöib trimetüülamiin moodustuda koliinist jmt. Imetajate jämesooles tekib metanooli pektiinist.
Ka atsetaat on hea allikas metaani tekkeks. Atsetaat lagundatakse klastilises reaktsioonis ja metüülrühma C redutseeritakse metaaniks ja karboksüülrühma C oksüdeeritakse CO2-ni. CH3COO- + H2O CH4 + HCO3- Atsetaadist suudavad metaani toota ainult vähesed metanogeenid nagu Methanosarcina jaMethanothrix (uus nimi Methanosaeta). Aga neid peab looduses olema suhteliselt arvukalt, sest on näidatud, et looduses moodustub 2/3 metaanist atsetaadist ja ainult 1/3 vesinikust ja CO2-st.
Atsetaadi baasil toimuv metanogenees on näiteks laialt levinud mageveekogude mudas, kus atsetaadi anaeroobset lagundamist teiste bakterite (anaeroobsed hingajad) poolt limiteerib teiste elektroni aktseptorite, nagu nitraadi ja sulfaadi defitsiit. Metanogenees on energeetiliselt vähe efektiivne. ATPd sünteesitakse prootongradiendi energia arvel ja ilmselt ainult 1 vöi veelgi vähem ATP-d tekib 1 mooli metaani tekkel.
Metaaniteke CO2-st, atsetaadist, metanoolist ja metüülitud ühenditest
CO2 fikseerimine metanogeenidel Kui metanogeenid kasvavad CO2 ja H2 peal, siis kasutavad nad CO2 ka C-allikana, on seega autotroofid. Kui nad aga kasutavad atsetaati või metanooli, siis kasutavad nad neid ühendeid ka C-allikana ja kui fikseerivad CO2, siis vähe.
Metanogeenid kasutavad CO2 sidumiseks reduktiivset mittetsüklilist rada (Ljungdahli-Woodi rada), kus CO2-st moodustub esmalt atsetüül-CoA ja siis sellest edasi teised C-ühendid. Atsetüül-CoA –st sünteesitakse pikema C-ahelaga ühendid, kasutades katkestatud TCA tsükli anaboolseid reaktsioone.
Reduktiivne AcCoA/ CO dh rada (Ljungdahli-Woodi rada) on lineaarne CO2 fikseerimise ahel, kus võtmepositsioonl on AcCoA, mis sünteesitakse kahest CO2 molekulist. Selline rada avastati esmalt atsetogeensetel klostriididel, kellel see osaleb atsetaadi tekkel CO2-st. Üks CO2 molekul redutseeritakse metüül-tetrahüdrometanopteriiniks nn metaanitekke rajas. Teine CO2 molekul redutseeritakse ensüümiga (CO DH) seotud karbonüülrühmaks ([CO]). Reaktsiooni katalüüsib CO dh. Sellesama ensüümi pinnal toimub ka komponentide kondenseerimine AcCoA-ks.
CO2 + 2e = CH3-H4MPT • CO2 + 2e = [CO] • [CO] + CH3-H4MPT + CoA= Ac-CoA Atsetogeensetel klostriididel asendab selles protsessis tetrahüdrometanopteridiini tetrahüdrofolaat
Üks CO2 molekul redutseeritakse metüülrühmani metaanirajas ja antakse üle CoB12-le. Teine CO2 redutseeritakse karbonüülrühmani CO DH pinnal. CO DH pinnal ühendatakse need kaks komponenti + CoA atsetüülCOA-ks
Methanosarcina Temal on heteropolüsahhariidne rakukest, mille koostis meenutab inimese sidekudede kondroitiini. See seob ka rakud agregaatideks. Kasutab metaani tootmiseks H2 + CO2, metanooli, atsetaati ja metüülamiini
Methanospirillumhungatii moodustab spiraalseid filamente ja spirillitaolisi rakke. Filamentide ümber on ühine tupp. Metaani moodustab formiaadist ning CO2 + H2-st.
Methanosaeta e. vana nimega Methanothrix moodustab niite. Toodab metaani atsetaadist
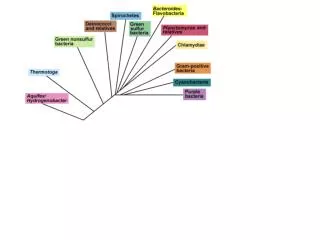
Archaeoglobus -mõneti sarnane metanogeenidega, aga ka krenarhedega Eurüarhed (sh metanogeenid) Krenarhed
Archaeoglobus- mõneti sarnane metanogeenidega, aga ka krenarhedega Archaeoglobus’e rakud on veidi ebaregulaarsed viburitega kokid, mis võivad olla üksikult või paarides. T opt on 83 kraadi. Agaril moodustavad rohekasmusti kolooniaid ja fluorestseeruvad 420 nm juures (sisaldavad koensüümi F420 nagu metanogeenidki). Nad on fakultatiivsed kemolitotroofid. Saavad oksüdeerida vesinikku ja tiosulfaati ning fikseerida CO2. Samas kasutavad ka orgaanilisi aineid, näiteks suhkruid. Kest on valguline.Redutseerivad sulfaate ja tiosulfaati, kuid mitte väävlit. On arhede hulgas ainus rühm, kus on sulfaatsed hingajad. Genoom sekveneeritud (ca 2400 geeni).
Ürgset tüüpi glükolüüs A. fulgidusel: ADP-seoselised kinaasid, Fd-seoseline GAP-i oksüdatsioon ja atsetaadi moodustumine Ac-CoAst üheetapiliselt.
Sulfaate redutseerivaid arhesid on seni leitud vaid süvamerre avanevatest vulkaanilistest allikatest. Põhjuseks võib olla see, et nad vajavad soola ja kõrget temperatuuri. Archaeoglobust saab rikastada anaeroobsetes tingimustes söötmes, mis sisaldab kunstlikku merevett ja sulfaati või tiosulfaati ja sobivat energiaallikat. T 85oC. C-allikana võib kasutada laktaati. Archaeoglobus oksüdeerib H2, laktaati, püruvaati, glükoosi jmt org. ühendeid, kandes elektronid sulfaadile või tiosulfaadile. Seega on ta anaeroobne hingaja.
Ferroglobus on sugulane Archaeoglobusega, kuid sulfaate ei redutseeri. Ta on rauda oksüdeeriv anaeroobnekemolitoautotroof, kes oksüdeerib nitraadiseoseliselt rauda. Ferroglobus võib energia saamiseks oksüdeerida ka H2S ja H2. Ta isoleeriti kuumast mereveest 85 oC juurest. Varem arvati, et raud hakkas Maal oksüdeeruma alles siis, kui tsüanobakteriaalse FS tulemusena moodustus suuremas koguses hapnikku. NB! Tegelikult saavad anaeroobsetes tingimustes rauda oksüdeerida ka mõned anoksügeensed FS bakterid.