Download

1 / 39
440 likes | 1.83k Views
FOXP2: the language gene. Talk of genetics and vice versa. Steven Pinker Commentary on Lai et al. The possibility that human language ability has genetic roots was raised about 40 years ago by the linguist Noam Chomsky and the neurologist Eric Lindberg.

E N D
Talk of genetics and vice versa • Steven Pinker • Commentary on Lai et al.
The possibility that human language ability has genetic roots was raised about 40 years ago by the linguist Noam Chomsky and the neurologist Eric Lindberg. • Chomsky noted that language is universal, complex, and rapidly acquired by children without explicit instruction. • Lindberg noted that a small number of children fail to display this ability, and that sometimes such deficits run in families.
Deficits of this kind are now called ‘specific language impairment’, an umbrella term for language disorders that cannot be attributed to retardation, autism, deafness, or other general causes. • Specific language impairment is more concordant in identical twins than in fraternal twins, indicating a heritable component. • The inheritance pattern is usually complex, and until recently poorly understood.
In 1990, researchers described the 3-generation KE family, half of whom have a speech and language disorder. • The disorder is distributed in the family in a way that suggests it is caused by a dominant gene on an autosomal (non-sex) chromosome. • Skeptics suggested that the gene merely lowers intelligence or makes speech unintelligible, or that the disorder is nothing more than an artifact of a working-class dialect.
Testing suggested that the disorder is more complex. • Affected family members do tend to score lower on average in IQ tests. • But the language impairment cannot be a simple consequence of low intelligence, because some of the affected members score in the normal range, and some score higher than their unaffected relatives.
Although the affected members have problems in articulating speech sounds and in controlled movements of the mouth and tongue, their language disorder cannot be reduced to a problem with motor control. • They also have trouble identifying basic speech sounds, understanding sentences, judging grammaticality, and other language skills. • For example, as adults they have difficulty with a task involving nonsense words that most 4-year-olds perform easily: completing sequences such as ‘Every day I plam; yesterday I ____’.
A forkhead-domain gene is mutated in a severe speech and language disorder. • Lai , C. S. L. et al.
Lai and colleagues studied the KE family in which a severe speech and language disorder is transmitted as an autosomal dominant single-gene trait. • Affected family members have a severe impairment in the selection and sequencing of fine orofacial movements, which are necessary for articulation. • The disorder is also characterized by deficits in several aspects of language processing, such as the ability to break up words into their constituent phonemes and grammatical skills including production and comprehension of word inflections and syntactical structure.
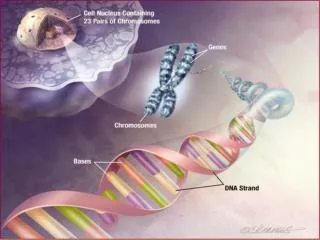
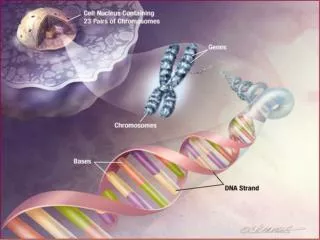
In previous work, they found that the gene mapped to a region 7q31 on chromosome 7. • Lai et al. found an individual CS, unrelated to the KE family, who had a very similar speech disorder. • When they examined the 7q31 region of CS, they found a chromosome translocation that disrupted a gene that they subsequently named FOXP2.
They went back to examine the FOXP2 in the KE family, they found a single mutation, a G-to-A SNP in exon 14, that was present in all affected individuals and absent in all unaffected individuals. • Lai et al. found that the mutation was absent in 364 chromosomes from unrelated Caucasian controls.
The FOXP2 gene is a member of the FOX family of transcription factors (genes that regulate the transcription of other genes). • The G-to-A SNP in exon 14 of FOXP2 is in a region that controls binding of FOXP2 to the DNA. It is invariant in all the currently known member of the FOX family, in species ranging from yeast to human.
Lai et al. suggest that the mutation observed in KE family members affects the DNA-binding or transcription properties of FOXP2. • FOXP2 is the first gene to have been implicated in the development of neural structures that are important for speech and language.
FOXP2 expression during brain development coincides with adult sites of pathology in a severe speech and language disorder. Lai , C. S. L. et al.
In vivo imaging technologies have developed that allow structural and functional neuroimaging. • MRI and PET revealed several abnormal brain structures in affected KE family members, compared with unaffected controls. • Lai et al decided to examine the expression of FOXP2 during brain development in mice and humans, and to compare it to abnormalities identified by imaging or suggested by regions known to be involved in language and motor control.
FOXP2 expression in the developing brain was not uniform or diffuse, but neither is it limited to any one brain area. • Instead, it shows restricted expression in a number of brain areas. • Many brain areas, such as the hippocampus, do not have detectable FOXP2.
FOXP2 is expressed in motor-related circuits during brain development. • In addition to expression in the developing cortical plate, FOXP2 expression during CNS development is found predominantly in a series of neural circuits that have been implicated in motor control, including the basal ganglia, thalamus, inferior olives, and the cerebellum. • These structures are intricately interconnected to subserve motor-related functions.
The basal ganglia modulate activity of premotor and prefrontal cortical areas via complex projections through the globus pallidus, substantia nigra, and the thalamus • The cerebellum plays an important role in regulating motor coordination, receiving input from the inferior olives. • Lai’s expression data implicating FOXP2 in the development of corticostriatal and olivocerebellar motor-related circuits during embryogenesis may account for the oromotor problems of humans with FOXP2 mutations.
It is possible that the accompanying linguistic and grammatical impairments in the KE family are secondary consequences of basic deficits in motor planning and sequencing. • However, it is equally plausible that the motor and cognitive problems arise simultaneously. • There is growing appreciation that area traditionally considered to be purely motor also contribute to cognitive and complex behavior.
The exclusively motoric nature of the caudate nucleus is challenged by data supporting roles in procedural learning and memory. • Similarly, it is now recognized that the cerebellum and prefrontal cortex form neural circuits with both motor and cognitive capabilities. • Thus, Lai’s data are consistent with the emerging view that subcortical structures play a significant role in linguistic functioning.
FOXP2 expression in sites of pathology identified by brain imaging • Lai’s observation of FOXP2 expression in the developing caudate nucleus of the embryo is paralleled by neuroimaging studies in the KE family. • A bilateral reduction of grey matter density in the caudate nucleus has been found in affected individuals. A PET study showed over-activation of the caudate nucleus in two affected individuals when performing a word repetition task.
FOXP2 mutation is also associated with significant structural abnormalities in the cerebellum, where Lai et al. found strong expression during embryonic development. • Studies of unrelated patients with acquired lesions highlight a cerebellar rle in procedural learning, particularly in detection and generation of event sequences and in linguistic functions.
FOXP2 expression patterns are highly concordant in mouse and human brain development • Lai et al. found a high degree of similarity between mouse and human FOXP2 expression patterns in the developing CNS. • They did not find any evidence for regions of FOXP2 expression that are only observed in humans in early brain development.
These data suggest that FOXP2 may be generally implicated in aspects of motor control in mammalian species, and was already playing a role in the development of motor-related brain regions in the human-mouse common ancestor. • Thus, positive selection of FOXP2 mutations in recent human history probably involved modifications to pre-existing brain systems, rather than acquisition of novel ones.
Molecular evolution of FOXP2, a gene involved in speech and language. • Enard W, Przeworski M, Fisher SE, Lai CS, Wiebe V, Kitano T, Monaco AP, Paabo S
The ability to develop articulate speech relies on capabilities, such as fine control of the larynx and mouth1, that are absent in chimpanzees and other great apes. • Enard et al. sequenced the complementary DNAs that encode the FOXP2 protein in the chimpanzee, gorilla, orang-utan, rhesus macaque and mouse, and compared them with the human cDNA. • They also investigated intraspecific variation of the human FOXP2 gene.
Enard et al. show that human FOXP2 contains changes in amino acid coding and a pattern of nucleotide polymorphism that strongly suggest that this gene has been the target of selection during recent human evolution. • FOXP2 (forkhead box P2) is located on human chromosome 7q31, and its major splice form encodes a protein of 715 amino acids belonging to the forkhead class of transcription factors2. • It contains a glutamine-rich region consisting of two adjacent polyglutamine tracts, encoded by mixtures of CAG and CAA repeats. • Such repeats are known to have elevated mutation rates.
In the case of FOXP2, the lengths of the polyglutamine stretches differed for all taxa studied. • Variation in the second polyglutamine tract has been observed in a small family affected with speech and language impairment, but this did not co-segregate with disorder, suggesting that minor changes in length may not significantly alter the function of the protein4. • If the polyglutamine stretches are disregarded, the human FOXP2 protein differs at only three amino-acid positions from its orthologue in the mouse.
When compared with a collection of 1,880 human–rodent gene pairs5, FOXP2 is among the 5% most-conserved proteins. • The chimpanzee, gorilla and rhesus macaque FOXP2 proteins are all identical to each other and carry only one difference from the mouse and two differences from the human protein, whereas the orangutan carries two differences from the mouse and three from humans. • Thus, although the FOXP2 protein is highly conserved, two of the three amino-acid differences between humans and mice occurred on the human lineage after the separation from the common ancestor with the chimpanzee.
These two amino-acid differences are both found in exon 7 of the FOXP2 gene and are a threonine-to-asparagine and an asparagine-to-serine change at positions 303 and 325, respectively. • Enard et al. compared the FOXP2 protein structures predicted by a variety of methods for humans, chimpanzees, orangutans and mice.
The chimpanzee and mouse structures were essentially identical and the orangutan showed only a minor change in secondary structure • The human-specific change at position 325 creates a potential target site for phosphorylation by protein kinase C together with a minor change in predicted secondary structure.
Several studies have shown that phosphorylation of forkhead transcription factors can be an important mechanism mediating transcriptional regulation. • Thus, although the FOXP2 protein is extremely conserved among mammals, it acquired two amino-acid changes on the human lineage, at least one of which may have functional consequences.
To investigate whether the amino acids encoded in exon 7 are polymorphic in humans, Enard et al. sequenced this exon from 44 human chromosomes originating from all major continents. • In no case was any amino-acid polymorphism found.
They analyzed the complete coding region of FOXP2 in 91 unrelated individuals of mainly European descent found no amino-acid replacements except for one case of an insertion of two glutamine codons in the second polyglutamine stretch4. • Because the two amino-acid variants specific to humans occur in 226 human chromosomes, this suggests that they are fixed among humans.
The evolutionary lineages leading to humans and mice diverged about 70 million years (Myr) ago. • Thus, during the roughly 130 Myr of evolution that separate the common ancestor of humans and chimpanzees from the mouse, a single amino-acid change occurred in the FOXP2 protein.
By contrast, since the human and chimpanzee lineages diverged about 4.6–6.2 Myr ago11, two fixed amino-acid changes occurred on the human lineage whereas none occurred on the chimpanzee and the other primate lineages, except for one change on the orang-utan lineage.
Enard et al. speculate that some human-specific feature of FOXP2, perhaps one or both of the amino-acid substitutions in exon 7, affect a person’s ability to control orofacial movements and thus to develop proficient spoken language. • If this speculation is true, then the time when such a FOXP2 variant became fixed in the human population may be pertinent with regard to the evolution of human language.
Under a model of a randomly mating population of constant size, the most likely date since the fixation of the beneficial allele is 0, with approximate 95% confidence intervals of 0 and 120,000 years. • Inclusion of population growth may push this time estimate back by at most the time since the onset of human population growth, some 10,000–100,000 years ago.
The method suggests that the fixation occurred during the last 200,000 years of human history, that is, concomitant with or subsequent to the emergence of anatomically modern humans. • This is compatible with a model in which the expansion of modern humans was driven by the appearance of a more-proficient spoken language.