Download
1 / 13
290 likes | 881 Views
Posttranslational modification is the chemical modification of a protein after its translation. Proteins often undergo several post-translational modification steps in parallel to protein folding. These modifications can be transient or permanent.

E N D
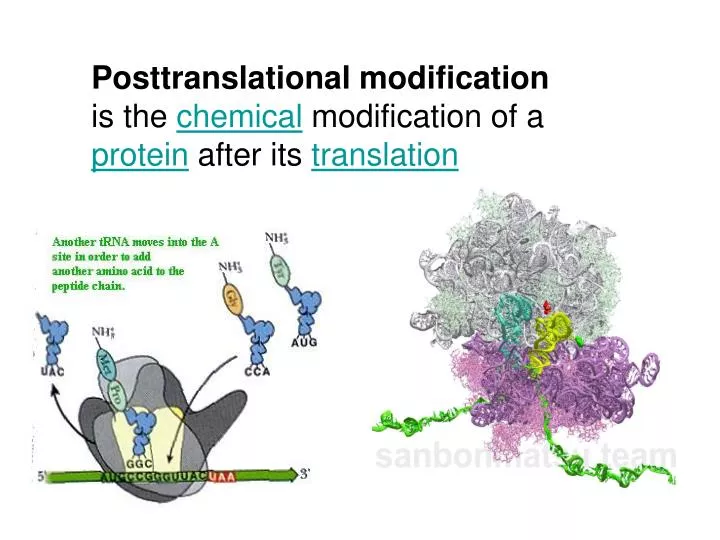
Posttranslational modification is the chemical modification of a protein after its translation
Proteins often undergo several post-translational modification steps in parallel to protein folding. These modifications can be transient or permanent. Most modifications are susceptible to alteration during the lifespan of proteins. Post-translational modifications generate variability in proteins that are far beyond that provided by the genetic code. Co- and post-translational modifications can convert the 20 specific codon-encoded amino acids into more than 100 variant amino acids with new properties. These, and a number of other modifications, can considerably increase the information content and functional repertoire of proteins
Post-Translational Modification 1. Dealing with the N-terminal residue In bacteria, the N-terminal residue of the newly-synthesized protein is modified to remove the formyl group. The N-terminal methionine may also be removed. In some cases the carboxy terminal residues are removed enzymatically 2. Loss of signal sequences • 15-30 residues at the amino terminal end of some proteins play a role in directing the protein to its ultimate destination in the cell. Signal sequences are ultimately removed by specific peptidases
Post-Translational Modification 3. Amino Acid Modifications • Many of the amino-acid side-chains can be modified Acetylation • The amino-terminal residues of some proteins are acetylated. This is more common in eukaryotes than prokaryotes e.g. the N-terminal serine of histone H4 is invariably acetylated as are a number of lysine residues.
Phosphorylation • In prokaryotes phosphorylation has been well established and shown to play an essential role in the control of bacterial protein function esp. enzyme activation. • E.g. the activity of isocitrate dehydrogenase is regulated via its reversible phosphorylation, which modulates the partition of carbon flux between the Krebs cycle and the glyoxylate bypass • Phosphorylation of proteins (at Ser, Thr, Tyr and His residues) is an important regulatory mechanism. For example, the activity of glycogen phosphorylase is regulated by phosphorylation of Serine 14. • Phosphorylation of tyrosine residues is an important aspect of signal transduction pathways. • Bacterial cells sense and respond to environmental signals through histidine phosphorylation
Methylation • In bacterial chemotaxis where in the absence of an added stimulus chemotaxis proteins are methylated to a basal level • Methylation of the 50S ribosomal proteins from Bacillus stearothermophilus, Bacillus subtilis, Alteromonas espejiana, and Halobacterium cutirubrum • The activity of histones can be modified by methylation. Lysine 20 of histone H4 can be mono- or di- methylated.
Carboxylation • Not very important in prokaryotes • The blood coagulation factor, prothrombin, contains a large number of carboxylated glutamatic acid residues in the N-terminal 32 amino acids. These modified residues are essential for activity. The modification requires vitamin K.
Hydroxylation • This does not occur in prokaryotes and can lead to problems with the expression of recombinant proteins • The conversion of proline to hydroxyproline in collagen is the classical example of a post-translational modification
Glycosylation • Numerous virulence factors of bacterial pathogens have been found to be covalently modified with carbohydrate residues, thereby identifying these factors as true glycoproteins. • In several bacterial species, gene clusters suggested to represent a general protein glycosylation system have been identified. • In other cases, genes encoding highly specific glycosyltransferases have been found to be directly linked with virulence genes. • Seems may be a role for glycosylation in pathogenesis. • both O-linked and N-linked protein glycosylation pathways in bacteria, particularly amongst mucosal-associated pathogens • Many extracellular (but not intracellular) proteins are glycosylated. Mono- or Oligo-saccharides can be attached to asparagine (N-linked) or to serine/threonine (O-linked) residues.
Nucleotidylation Mononucleotide addition is used to regulate the activity of some enzymes. Two different examples are found among the system that regulates Nitrogen utilization in E. coli: • Glutamine synthetase is adenylylated (i.e. AMP is added) at a specific tyrosine residue. The enzyme is inactive when it is adenylylated. The degree of adenylylation is controlled by a regulatory protein, PII. • The ability of PII to regulate the adenylylation of glutamine synthetase is in turn regulated by its own uridylylation (i.e. the covalent addition of UMP). PII is also uridylylated at a tyrosine residue.
4. Lipid Addition • Some proteins have lipid moieties attached: • The viral src protein is myristoylated at the N-terminal glycine. • Rhodoposin is palmitoylated at a cysteine residue • The ras oncogene protein is farnesylated as are some G proteins. • Some eukaryotes, notably parasitic protozoa, have glycosylphosphatidylinositol-linked proteins. 5. Adding Prosthetic Groups • Proteins that require a prosthetic group for activity must have this group added. For example, the haem (heme) group must be added to globins and cytochromes; Fe-S clusters must be added to ferredoxins. 6. Forming Disulfide Bonds • Many extracellular proteins contain disulfide cross-links (intracellular proteins almost never do). The cross-links can only be established after the protein has folded up into the correct shape. • The formation of disulfide bonds is aided by the enzyme protein disulfide isomerase in eukaryotes and by the DsbA protein in bacteria.
7. Proteolytic Processing • Some proteins are synthesized as inactive precursor polypeptides which become activated only after proteolytic cleavage of the precursor polypeptide chain. E.g. Chymotrypsin & Trypsin • These are both synthesized as zymogens. • Cleavage of chymotrypsinogen between Arg15 and Ile 16 by trypsin yields the enzymatically active pi-chymotrypsin. • Two further proteolytic cleavages catalyzed by chymotrypsin removes the dipeptides Ser14-Arg15 and Thr147-Asn148 to yield alpha-chymotrypsin.






















