Download

1 / 1
10 likes | 132 Views
B. A. ANNEALING STUDIES OF MULTI-DIMENSIONAL DNA ORIGAMI. C. D. Tanner Bakhshi, Molly McIlvain , David Neff, and Michael Norton, Ph.D.: Department of Chemistry, Marshall University. ABSTRACT.

E N D
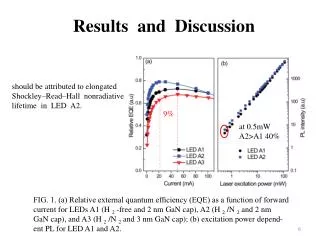
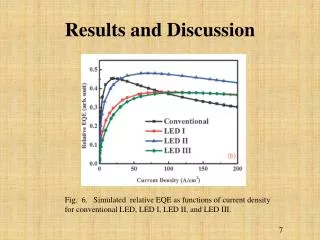
B. A. ANNEALING STUDIES OF MULTI-DIMENSIONAL DNA ORIGAMI C. D. Tanner Bakhshi, Molly McIlvain, David Neff, and Michael Norton, Ph.D.: Department of Chemistry, Marshall University. ABSTRACT In 2006 Paul W. K. Rothemund of Caltech published an article in Nature about something he called “DNA Origami,”1a technique involving the manipulation or folding of DNA to form virtually any desired shape or pattern. In the seven years following this discovery, DNA Origami has become a sensational research topic in many disciplines, namely the field of DNA nanotechnology. With the many potential benefits of this technology, however, come many challenges. One challenge in particular is the length of time that it can take to anneal a long DNA strand to short “staple strands” that “show” the longer strand of DNA how and where to fold. Standard protocol for this procedure calls for an eighteen-hour temperature ramp using a Polymerase Chain Reaction (PCR) machine. By staining the DNA with a fluorescent nucleic acid stain (SYBR Green I, Cambrex), it is possible to obtain fluorescent intensity data while annealing DNA origami. As more and larger DNA structures begin to form, fluorescence increases proportionally, thus providing a map that highlights temperatures that are crucial in origami formation. After analyzing this data, it was evident that multiple temperature ranges were not instrumental in the formation of origami. It was hypothesized that if a temperature ramp protocol including only those ranges that appeared to be critically important in origami formation were developed, then origami could be annealed in a significantly shorter amount of time, a mere thirty minutes. Results showed that the thirty-minute protocol was capable of forming origami after only this short annealing time. Using atomic force microscopy (AFM), samples of origami from both protocols were characterized and compared. The samples were determined to be equally sound in form and structure. The shorter, thirty-minute method of origami formation can be used as a viable alternative to the eighteen-hour procedure when fast experimental turnaround is desired. INTRODUCTION RESULTS AND DISCUSSION Materials: Each attempt to shorten origami production time began by mixing individual solutions of 0D, 1D, and/or 2D origami [IDT (staple strands), Bayou Biolabs (7249-base single-stranded M13mp18 DNA) in Tris-Acetate-EDTA/MgCl2 buffer, pH 8, SYBR Green 1, Cambrex]. Solutions were mixed and stored in duplicates in plastic tubes (Applied Biosystems) connected in rows of eight. The eighteen-hour annealing method, as previously mentioned, was run on a real-time PCR machine (Model 7300, Applied Biosystems), and the faster program was run in a thermal cycler (Model Primus 96 Plus, MWG-Biotech). For this experiment programmed temperature ramps always began at the highest temperature and ended at the lowest temperature. In 2006 Paul W. K. Rothemund, a researcher at the California Institute of Technology, published an article in Nature about something he called “DNA Origami.”1True to the way that it sounds, DNA Origami involves the folding of DNA into different patterns and shapes through the use of “staple strands” that tell the DNA where and how to fold. The type of origami used in this experiment is called “cross” origami.2Just like it sounds, the shape is formed by the joining of two linked but individually- formed rectangular DNA structures, the end result’s being what can be described as a cross or plus sign. Depending on how the ends of these separate structures are designed, the crosses formed can be individual, they can stick together in one direction and form long chains, or they can stick together in two directions and form large checkerboard-like arrays. These different configurations are referred to as 0D, 1D, and 2D crosses, respectively. A. B. Though DNA Origami has become a sensational research topic in many disciplines and poses many potential benefits, studying it does not come without challenges. One challenge in particular is the length of time that it can take to anneal a long DNA strand to the aforementioned staple strands. Standard protocol for this procedure calls for an eighteen-hour temperature ramp. It was hypothesized that if a temperature ramp protocol including only those ranges that appeared to be critically important in origami formation were developed, then origami could be annealed in a significantly shorter amount of time, a mere thirty minutes. Fig. 3. AFM images of (A) 1D cross origami produced using the traditional 18- hour anneal, (B) 2D cross origami produced using the traditional 18-hour anneal, (C) 1D cross origami produced using the rapid 30-minute anneal, and (D) 2D cross origami produced using the rapid 30-minute anneal. Fig. 3 A and B show 1D and 2D cross origami, respectively. These origami were annealed using the eighteen-hourmethod in the real-time thermal controller. The 1D origami, though tightly clustered, was well formed (The “equal signs” discussed in Fig. 1 are present.), and the chains had typical lengths of ~1µm, as we have seen in previous experiments. The 2D origami also appeared to have formed adequately and were arranged in fairly large arrays. Fig. 3 C and D, too, show 1D and 2D cross origami, respectively. This time, though, the origami were annealed using the thirty-minute thermal-cycler method instead. The 1D origami, like that of the eighteen-hour run, had precise form (“equal signs”). However, the chains appeared to be longer (> 1µm) than those featured in Fig. 4 A. (Considering that each cross is only about 100 nm in width, a 1.7-micrometer chain is huge!) The 2D origami was especially pleasing, featuring what looked to be a very solid 6x6 cross array (“equal signs”). In only thirty minutes, we were able to produce origami that was qualitatively verysimilar to origami produced over an eighteen-hour period. This allows for quick turnaround during future experimentswhich is necessary when future plans depend upon the answers that may lie within a present experiment. The results of this experiment, and the prospect of a quick alternative to a conventional method, are encouraging. EXPERIMENTAL C. Method: The first step in producing DNA Origami on a shorter time scale was to gather information about its formation over the longer, eighteen-hour time period using a real-time PCR machine. In addition to providing the environment necessary for origami growth, the machine also monitored the intensity of light produced by fluorescent materials. In the presence of double-stranded DNA, SYBR Green 1 (SG1) shows increased fluorescent activity relative to that seen in the presence of single-stranded DNA. Therefore, as more, larger DNA structures begin to grow, the fluorescence of the indicator increases proportionally. Thus, as the thermal controller (real-time PCR machine) monitored fluorescence emissions data and sent a digital readout to the computer operating it, the behavior of DNA Origami could be seen graphically. If derivatives of these numerical values of fluorescence were calculatedand the resulting values graphed as a function of temperature, then the growth activity of origami could be seen on a temperature-by-temperature basis, and data could be treated more or less like a histogram showing annealing activity.3 It became apparent when studying the graph that certain temperature ranges, both high and low, indicated little to no origami formation. These temperature ranges were ignored, and focus was shifted instead to those ranges where change in fluorescent activity (ΔF) over time(ΔT) was highest. Derivatives were taken of the numerical ΔF/ΔT values obtained from the thermal controller for all important temperatures (Figs. 2A and 2C), and these derivative values were then summed. By dividing each temperature’s fluorescence value by the sum total of fluorescence, each temperature was assigned a percentage. These percentages provided a quantitative representation of the amount of change in SYBR Green I fluorescence that occurred during the eighteen-hour period. Thus, it was possible to determine which temperatures were the most effective in annealing staple strands to M13 DNA strands, i.e. forming origami. These percentages were multiplied by 1800 (the number of seconds in thirty minutes), revealing the length of time that origami solutions should remain at each temperature during the thirty-minute program (Figs. 2B and 2D). Cross origami annealed using thirty-minute programs were then characterized using atomic force microscopy, followed by a comparison to eighteen-hour cross origami. D. LITERATURE CITED 1) Rothemund, Paul W. K. Folding DNA to Create Nanoscale Shapes and Patterns. Nature. 2006, 440, 297-302. 2) Liu, Wenyan; Zhong, Hong; Wang, Risheng; Seeman, Nadrian C. Crystalline Two-Dimensional DNA-Origami Arrays. Angewandte Chemie International Edition. 2011, 50, 264-267. 3) Arbona, Jean-Michel; Elezgaray, Juan; Aime, Jean-Pierre. Modelling the Folding of DNA Origami. Europhysics Letters. 2012, 100, 28006. ACKNOWLEDGEMENTS Fig. 2. Graphs showing (A) the derivatives (ΔF/ΔT) taken of the numerical values of intensity at each temperature during 1D cross origami formation, (B) the amount of time spent at each temperature in the thirty-minute program for 1D cross origami, (C) the derivatives (ΔF/ΔT) taken of the numerical values of intensity at each temperature during 2D cross origami formation, and (D) the amount of time spent at each temperature in the thirty-minute program for 2D cross origami formation. Note that annealing programs (B and D) were made after derivative data was gathered and analyzed (A and C). • Funding for this project was provided by the U.S. Army through the High School Apprenticeship Program (HSAP). • Tanner Bakhshi would like to thank Dr. Michael Norton, David Neff, and Dr. Masudur Rahman for guidance and support.