Download

1 / 46
460 likes | 676 Views
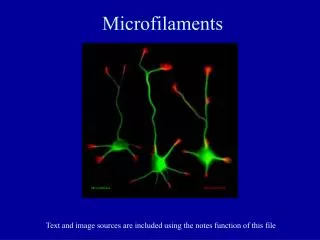
Dissecting the Complexity of Cell Structure 2: Microfilaments. How actin-based polymers contribute to cellular structure, behavior, and motility. An overview of the actin cytoskeleton in a cultured epithelial cell, using antibodies and immuno- fluorescence.

E N D
Dissecting the Complexity of Cell Structure 2: Microfilaments How actin-based polymers contribute to cellular structure, behavior, and motility
An overview of the actin cytoskeleton in a cultured epithelial cell, using antibodies and immuno- fluorescence. Actin is at the cell cortex and also organized into fibers that extend through the cell.
Formation of microfilaments (MFs) from the protein actin
Actin polymerization is controlled, in part, by an initiating complex that includes actin-like proteins
The Arp2/3 complex not only initiates MF polymerization, it can bind to the walls of existing MFs, promoting the formation of branches, which turn that region of the cell into a gel (as opposed to a “sol”).
Actin at the edge of a cultured cell, looking at all actin (top) or only recently added actin (bottom). Actin polymerizes at the cell periphery
Direct evidence for the addition of actin monomers to the membrane-end of MFs
Actin polymerization is also regulated by small proteins that bind to soluble actin and modify its behavior in solution. Here is profilin, which catalyzes the exchange of ADP for ATP in the actin cleft, increasing the pool of polymer- ization-competent monomer
There are also proteins that retard MF growth: While profilin enhances MF growth by increasing the concentration of ATP-actin, Thymosin reduces it by making a complex that will not polymerize
Diagramatic representation of the pathways The help to regulate MF formation
So how does a complex process like this actually work? • Amoeboid motility does not require MTs: it proceeds normally in the presence of MT poisons • It is immediately poisoned, however, by drugs that block actin polymerization, like Cytochalesins D and E or Latrunculin A • It fails in several mutant strains that lack key actin assembly proteins (though it is amazingly robust to mutation) • Infer, to understand amoeboid movement and related kinds of cell motion we need to understand the control of actin polymerization and organization
Proteins that bind to the walls of MFs can bind them together in different geometries
Four proteins that bind the sides of MFs and bundle them into different geometries
TEM image and diagram of MF Bundling in a microvillus
Diagram of MF web formed when cross-linking is done by the long, fibrous protein, Filamin
When filamin is under-expressed, due to mutation, cell morphology and motility are abnormal. Cells depleted for Cell expressingfilamin by LOF mutation normal filamin levels
Like MTs, MFs can bind some associated proteins that alter the properties of the polymer. Cofilin binds to F-actin and distorts the polymer, making it less stable.
Gelsolin also bind the MF wall, distorting is and inducing breaks that shorten the average fiber.
Gelsolin and proteins like it can break up MF either in vitro (as shown here) or in vivo, leading to rapid reshaping of the actin cytoskeleton
Breaks in MFs mean both more and shorter MFs therefore more ends (for a given amount of polymer). Thus, both growth and shrinkage of polymer can be faster after MF severing.
Capping proteins, like Cap-Z can silence one MF end for further subunit addition; in this case it is the fast-growing (plus) end that is turned off.
All these processes can be regulated to make For a quite complex behavior of the MFs in vivo. Blood platelets (thrombocytes) looking as they do in circulating blood (left), after attachment to a surface (center) and during a platelet “reaction”, where they attach to a substratum and contract. All this is MF assembly-dependent.
But What Initiates MF Polymerization at the PM?
Projection morphology and other actin-dependent shape changes suggest that controlled polymerization can also give rise to unidirectional fibers, the filopodia • Microvilli are of this form • Dynamic projections from not only platelets but also neuronal growth cones are of this form • Even projections from sperm and egg, involved in fertilization, are of this form • There must be a membrane-associated actin initiator that does not involve branching
Formins are now recognized as molecules that can initiate MF polymerization at the PM and allow continued addition of actin at the PM while an association with the membrane is maintained • Formins contain conserved domains, the Formin-homology (FH) domains. FH1 and 2 define a comparatively short polypeptide that can serve as an efficient nucleator of MF polymerization • Formin-initiated polymerization adds actin subunits AT the PM, pushing the already formed MF away • This allows PM-controlled events to regulate aspects of MF organization, much as MTOCs control MT formation
Movements of PM-associated “cortical” actin can affect the position and organization of other components of cytoplasm, like MTs
Conversely, the behavior of MTs can affect behavior of the actin cortex
Actin “stress fibers” are initiated by and attached to the cell cortex.Cell margin seen with The same region seen withoptics that show proximity fluorescence optics and anbetween the cell’s surface antibody that lights up actin and the underlying substratum These are “focal adhestions” that mark site of cell attachment
Another view of stress fibers (green) and “focal adhesions”, stained with antibodies to vinculin, a component of stress fiber adhesion sites
There are specific trans-membrane proteins and adaptor proteins that connect MFs with the extracellular milieu
To promote tissue strength, cadherins are specifically but non-covalently bound to the MFs of the cytoskeleton. The linking proteins include catenins, which play a role in signaling between tissues.
Cadherins bind the MF cytoskeleton of one cell to that of its neighbors, forming a mechanical unit. This coupling contributes to the mechanical integrity of a tissue.
In some cells, cadherins and actin MFs form ordered arrays that can work like a contractile ring and control the cell’s diameter.
Integrins are membrane proteins that bind cells to the “Extra-Cellular Matrix” (ECM). The integrins make bonds between the actin cytoskeleton and the fibers of the ECM, such as collagen and fibronectin.
Transmembrane proteins link the actin-dependent cytoplasmic motility that pulls on MTs with extracellular material, such as beads that can attach to the cell’s surface
Thus, we can recognize a mechanical continuum that runs from cytoplasmic MFs, controlled in part by MTs, through the plasma membrane to the molecules of the extracellular matrix (ECM). • The questions defined by this idea include the identity and behavior of the linker molecules (cadherins, integrins, and others) and the processes and molecules that control the behavior of the fibers and their states of polymerization and linkage
Signal transduction cascades that activate small G-proteins regulate MFs in a wide range of cells
Reminder of the mechanism for signaling by small G-proteins, like Rac, Rho, and CDC-42: GTP-bound state is active and turns on down-stream proteins. GAPs and GEFs control the relative concentrations of GTP- and GDP-bound forms of the protein
In budding yeast there is a signaling cascade that links a pheremone to regulation of the actin cytoskeleton Discovery of such control pathways has motivated a Search for homologous paths in other cell types
There are >8 down-stream targets for each of the small G-proteins involved in regulation of the actin cytoskeleton • Rho-GTP leads to the activation of cytoplasmic myosin, which in turn promotes actomyosin-dependent structures, like stress-fibers • Rac-GTP activates WASp, which in turn activates the Arp2/3 complex, and it also promotes uncapping of MFs by removing CapZ or Gelsolin, inducing extensive, branched +-end MF growth • CDC42-GTP also activates the WASp protein but Formin is also activated, promoting MF assembly into membrane-associated bundles
A plausible mechanism for the organization of actin in the cell’s cortex and for its role in extension of a lamellipod
Signal transduction cascades that activate small G-proteins have now been found in a wide range of cells
Functional Rac is important for mebrane-associated actin’s response to MT invasion of the cell cortex
Compare cortex behavior with a cell expressing wild-type Rac during MT regrowth