Download

1 / 50
510 likes | 750 Views
Centrální dogma molekulární biologie. Alexandr Sember. Genetika (2009) český překlad originálu Principles of Genetics 5th edition (Snustad a Simmons, 2009). Molecular Biology of the Gene 6th edition (2008) J. D. Watson et al. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.TOC&depth=2.

E N D
Centrální dogma molekulární biologie Alexandr Sember
Genetika (2009) český překlad originálu Principles of Genetics 5th edition (Snustad a Simmons, 2009) Molecular Biology of the Gene 6th edition (2008) J. D. Watson et al.
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.TOC&depth=2http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.TOC&depth=2 http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.TOC&depth=2
malé mol. ionty 4% fosfolip. 2% DNA 1% nukleové kys. 30% ostatní RNA 6% 70% voda proteiny 15% polysach. 2% Složení buňky • cca. 15% celkové hmotnosti buňky (55% sušiny) tvoří proteiny • Buňka má informaci o syntéze proteinů uloženou v DNA
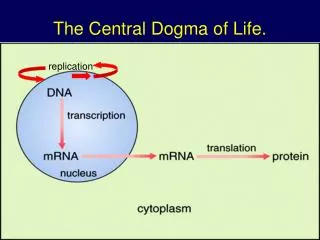
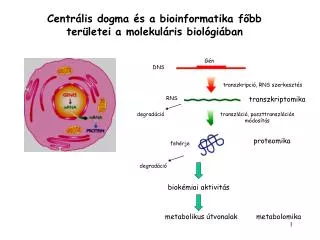
Centrální dogma MB • Francis Crick (1956) • Směr přenosu genetické informace: chromozomální DNA funguje jako templát pro RNA, která se poté přemístí do cytoplasmy, kde funguje jako templát pro proteosyntézu • základní myšlenka stále platná, pouze později doplněna o možnost reverzní transkripce (Temin, Baltimore, 1970 – objevení reverzní transkriptázy u viru Rousova sarkomu, 1975 NC); pochopitelně je i možné přepsat RNA do další RNA
DNA jako genetický materiál • 1868 – Johann Friedrich Miescher • Izolace nukleinu z buněčných jader • obvazy z gangrén nasáklé hnisem (leukocyty) • Hrubé extrakty DNA (několik desítek bp) • Unikátní poměr P/N, chybí S • 1869 – 1940 – Phoebus Aaron Theodor Levene • - pracoval na složení NK • zjistil, že DNA má 4 základní kameny (báze) • Naměřil,že jejich poměr je 1:1:1:1 • tetranukleotidová hypotéza – pořadí nukleotidů je neměnné, tetranukleotid se pravidelně opakuje, proto nemůže vyjadřovat žádnou genetickou informaci; NK je lešení, na němž jsou navěšené proteiny; chemickou podstatu genu je třeba hledat ve struktuře proteinů
DNA jako genetický materiál 1928 – Frederick Griffith - pokus se Streptococcus pneumoniae (zápaly plic u člověka; smrt u myší) - nebezpečnost souvisí s obalením do polysacharidového pouzdra S-kmen - aktivní gen, který kóduje enzym pro syntézu kapsulárního polysacharidu - hladký vzhled (S = smooth, hladký) R-kmen (R= rough, drsný vzhled), nemají pouzdro a nejsou proto virulentní
DNA jako genetický materiál Griffithův experiment – došlo k přenosu genetického materiálu mezi dvěma kmeny bakterií procesem transformace 1. virulentní S - kmen způsobí smrt myši; 2. nevirulentní R – kmen, myš přežije; 3. – mrtvé virulentní kmeny – nic se neděje; 4. – mrtvé virulentní kmeny smíchané s živými nevirulentními → myš zemřela; izolace živých virulentních kmenů
DNA jako genetický materiál 1944 – Avery, McLeod, McCarty • Transformační princip – komponenta mrtvé bakterie zodpovědné za transformaci • Opakování Griffithova experimentu • Využití rozkvětu enzymologie (proteázy, RNázy, DNázy) • Pouze při působení DNáz experiment přestal fungovat • Definitivní důkaz - DNA = gen. materiál • 1941 – William Atsbury • jako první izoloval opravdu vysokomolekulární DNA • první difrakční obrazce • 1950 Erwin Chargaff • množství všech 4 bází není ekvimolární • relativní podíl bází není náhodný (A=T, C=G) • AT se nerovná GC, zastoupení u různých organismů se liší = vyvrácení tetranukleotidové hypotézy • Chargaffovo pravidlo: poměr pur/pyr se rovná vždy 1 nezávisle na tom, z jakého zdroje DNA pochází
DNA jako genetický materiál • Chargaffovo pravidlo: obsah pyrimidinů = obsah purinů
DNA jako genetický materiál • 1952 – Alfred Day Hershey, Martha Chase (NC) • Je DNA genetickým materiálem i u virů? • analýza různých složek bakteriofága T2 • využití pokroku (radioizotopy, izolace fágů, mikrobiologie E. coli) • Kapsidy fágů zůstávají na povrchu x genetický materiál jde do bakterie
kultura E. coli s bakteriofágy → kultivace na médiu s radioaktivním P+S • S → chybí v DNA • P → chybí v proteinech (výjimky: fosforylace) 2 experimenty – v jednom případě značena pouze DNA a ve druhém pouze proteiny • značené proteiny virového obalu zůstaly mimo buňku • značená DNA viru se dostávala do buňky + vznik nového fágového potomstva • definitivní důkaz, že za přenos dědičné informace je zodpovědná DNA i u virů • RNA jako genetický materiál byla objevena roku 1957 u viru tabákové mozaiky TMV (Fraenkel-Conrat + Singer)
Rentgenová difrakční analýza William Atsbury, Maurice Wilkins, Rosalind Franklin Rentgenová difrakční analýza – molekuly měřené struktury dokážou vychýlit rentgenový paprsek v závislosti na struktuře a molekulární hmotnosti (typu atomů) 1951 – Rosalind Franklinová – nejlepší difrakční obrazce
1953 – James Watson, Francis Crick • trojrozměrný model struktury DNA • Syntéza z cizích výsledků (Franklin, Chargaff..) • Důležitý ne přímo ten model, ale hlavně návrh, jak by se DNA mohla replikovat!! • 1962 Nobelova cena za fyziologii a lékařství (spolu s Wilkinsem, ale bez Franklinové – zemřela ve 37 letech na rakovinu vaječníků, NC se neuděluje in memoriam)
kříž = šroubovice chybějící 4. řada pruhů ukazuje, že DNA je double helix Báze vrstveny kolmo k ose Vzdálenost mezi bázemi = 3,4 Å 1 otočka = 34 Å = 10 bp (ve skutečnosti 10,4) Průměr šroubovice = 20 nm 1 nm = 10-9 metru 1 Ångström (Å) = 0,1 nm
Struktura NK Primární struktura = posloupnost nukleosidtrifosfátů (NTP, dNTP) pospojovaných fosfodiesterovou vazbou (resp. pořadí bází);Nevětvené lineární molekuly Nukleotid v se skládá z: 1) Sacharidová jednotka (cukr, furanóza) DNA: 2-deoxy-β-D-ribóza RNA: β-D-ribóza 2) Dusíkatá baze – adenin, guanin, thymin, cytosin, uracil Purinové baze: A, G Pyrimidinové baze: C, T, U 3) Zbytek kyseliny trihydrogenfosforečné Nukleotid = nukleosid (cukr+báze) + fosfát
Struktura NK Nukleotid v se skládá z: 1) Sacharidová jednotka (cukr, furanóza) DNA: 2-deoxy-β-D-ribóza RNA: β-D-ribóza 2) Dusíkatá baze – adenin, guanin, thymin, cytosin, uracil Purinové baze: A, G Pyrimidinové baze: C, T, U 3) Zbytek kyseliny trihydrogenfosforečné N-glykosidická vazba Pozice na cukru se označují tzv. „s čarou“ (na bázích ne) Největší rozdíl mezi DNA a RNA je 2´OH skupina RNA
Struktura NK Nukleotidy: adenosin guanosin thymidin mono-di-tri -fosfát cytidin uridin AMP – ADP – ATP dATP x ATP
Struktura NK Puriny: aromatické heterocyklické báze, rovinný (= planární) dvojcyklus, v poloze 9 je navázán cukr Adenin = 6-aminopurin (analog je mutagenní 2-aminopurin) Guanin = 2-amino-6-ketopurin
Struktura NK Pyrimidiny: aromat. heterocykly, jednoduchý cyklus, v poloze 1 je navázán cukr Cytosin = 2-keto- 4-aminopyrimidin (O= na C2 a NH2 na C4) Thymin = 2,4 – diketo – 5 – metylpyrimidin (O =na C2 a C4, metyl na C5) Uracil = 2,4 – diketopyrimidin (O= na C2 a C4) Thymin je vlastně 5-metyluracil Deaminací cytosinu vzniká uracil Deaminací 5-metylcytosinu vzniká thymin Zapamatujte si uracil a máte vyhráno
Tautomerismus bází Přeskakování vodíku mění uspořádání donor/akceptor vodíkové vazby – jiné preference párování Stabilní formy bází = keto (T, G, U), amino (A, C) Méně stabilní izoformy = enol (T, G, U), imino (A, C)
Zdroj metylu pro metylace Nukleotidy mohou mít i jiné funkce… ATP = zdroj energie v živých systémech Kofaktory enzymů Kofaktor enzymů Druzí poslové v signalizaci
Sekundární struktura DNA = 2 molekuly ssDNA spojené vodíkovými vazbami (Watson-Crickovské párování bází) Báze jsou komplementární (donor/akceptor vodíkové vazby) A-T 2 vodíkové vazby G-C 3 vodíkové vazby Páry AT a GC jsou stejně široké (překryv C1 deoxyribózy = N-glykosidická vazba, symetrie helixu) Vodíková vazba = interakce mezi silně elektronegativním atomem (O, F, N) a vodíkem NH2 = donor H-vazby =O, N = akceptor H-vazby
28 typů párování nukleotidů volně v roztoku př. Hoogsteenovo párování – rekombinace, G-kvartet; wobble párování – využití při čtení genetického kódu
Polarita řetězců dána fosfodiesterovou vazbou 5´ ATTGCCA 3´ 3´ TAACGGT 5´ 5´-P a 3´OH Watson-Crickovské párování Antiparalelní uspořádání Pravotočivá dvojšroubovice Báze jsou uvnitř, cukr-fosfátová kostra vně Malý a velký žlábek Sekundární struktura DNA = pravotočivá/levotočivá dvojšroubovice Terciální struktura DNA= lineární, kružnicová, nadšroubovice Kvarterní struktura DNA = pouze ve smyslu nějakého komplexu s proteiny
Malý a velký žlábek Důvody: páry bází neprocházejí přesně středem podélné osy dvoušroubovice N-glykosidické vazby nevycházející z páru bází pod úhlem 180°. Úhly mezi oběma vazbami jsou 120° (malý žlábek) a 270° (velký žlábek) 270° • Důsledek: • Jiná nabídka chemických skupin • v malém žlábku chybí metyl – rozpoznání pouze páru AT nebo GC • Ve velkém žlábku rozpoznání konkrétní báze (využití regulačními proteiny, regulace transkripce x TBP vazba do malého žlábku) 120°
Stabilita DNA • 1) Stacking interakce • hlavní příspěvek ke stabilitě DNA • Vrstvení bází na sebe (hydrofobní a van der Waalsovy interakce) • splývání oblastí výskytu delokalizovaných π-elektronů sousedících aromatických kruhů • GC páry mají mnohonásobně silnější stacking interakce mezi sebou (než AT páry) 2) Hydratační obal • Primární (pouze voda) a sekundární (+ ionty) • Primární obal: 20 molekul vody / 1 nt • Zeslabení odpuzování sousedních fosfátů • Hydrofobní interakce (posiluje stacking) • Doplnění dalších vodíkových můstků • 3) Vodíkové vazby mezi bázemi • Velmi malý příspěvek ke stabilitě • Význam spíše pro specifitu párování
Konformace DNA/RNA • Konformační polymorfismus • Různá hydratace a koncentrace solí • Modifikace bází (př. 5-metylcytosin u Eukaryot) • Alternativní párování bází, vrtulovité zkroucení bází apod. • Konformační flexibilita ribózy a N-glykosidické vazby • Sekvenční polymorfismus – závislost struktury na lokální sekvenci
3D struktura molekuly: Popis torzními (dihedrálními) úhly – jejich součet = 180° 0° = syn 180° = anti Máme atomy ABCD, které jdou ve vazbách za sebou; torzní úhel je úhel mezi vazbami AB a CD podle vazby BC, která tvoří jakoby pant mezi nimi
N – glykosidická vazba • Všechny nukleotidy preferují anti • Pouze G preferuje aspoň z 15% syn • Konformace ribózy • Pokud jsou C4, O4 a C1 v jedné rovině, sledujeme, co je nad a pod rovinou cyklu • V DNA obvykle C2-endo, fosfáty pohodlně daleko od sebe (70 Å) • V RNA a hybridech DNA/RNA obvykle C3 – endo (kvůli 2´OH skupině)
Konformace DNA/RNA • Ribóza ve C3-endo • RNA, hybrid DNA/RNA • Báze vytlačeny od středu, uprostřed kanál • C2-endo, klasická forma DNA • Žlábky pěkně vykreslené • C2-endo, G v syn • Schodovitá struktura, opakování dvou párů bází • Levotočivá!!
Elektrické vlastnosti NK • záporný náboj, pohyb v elektrickém poli • chovají se jako polyanionty (dsDNA má dva mínus náboje/1bp) • separace na agarózovém nebo PAA gelu = elektroforéza (anionty jako např. NK jdou k kladné elektrodě, tedy k anodě), vizualizace např. ethidiumbromidem (interkalační činidlo) • dělení podle velikosti a topologie, menší molekuly procházejí rychleji
Denaturace NK • Reverzibilní narozdíl od proteinů • Denaturace teplotou, pH (pod 3, nad 11), formamidem (interference H-můstků), močovinou, enzymaticky (helikáza) • Denaturace a reasociace (repetitivní sekvence rychleji) • Stabilita duplexů klesá v řadě RNA/RNA DNA/RNA DNA/DNA • Tání duplexů lze sledovat pomocí měření absorbance v UV-oblasti • NK mají absorpční maximum při vlnové délce 260 nm, proteiny při 280 nm • Tm = teplota tání = teplota, při které je polovina duplexu denaturována; je tím vyšší, čím je větší obsah GC • Hyperchromní efekt = zvýšení absorbance i přesto, že koncentrace DNA se nemění (při denaturaci volné báze lépe absorbují UV) • dsDNA má absorbanci o 40% menší než dvě ssDNA
cílová DNA denaturace hybridizace sonda Hybridizace FISH = Fluorescence in situ hybridization X Southern, Northern blot = hybridizace in vitro
Rozdílné vlastnosti DNA/RNA v extrémním pH Silně kyselé pH DNA: bude snáze docházet k depurinacím, DNA zdenaturuje, ale i zdegraduje RNA: pouze zdenaturuje (stabilní při pH 4 – 5,2) Silně zásadité pH DNA: bude pouze denaturovat RNA: deprotonace 2´OH skupiny – nukleofilní atak fosfodiesterové vazby– vznikne 2´- 3´cyklický fosfát - rozpad na směs 3´ +2´ monofosfátů = denaturace+degradace
Struktura RNA • Vyšší flexibilita, strukturní a funkční mnohotvárnost • Často ssRNA, menší velikost • Rozmanité sekundární struktury (vlásenky, bubliny…) • Terciální struktury (pseudozel..) 16S rRNA • Stabilita: intramolekulární interakce (za přispění 2´OH a nekonvenčního párování pází), stacking interakce, vazba dvojmocných kationtů, stabilní tetraloops (rRNA), mnoho modifikovaných bází a případná metylace 2´OH, tautomerie bází
Využití 2°struktury (vlásenka, SECIS) k inkorporaci selenocysteinu do proteinu Využití 3°struktury (4 pseudouzly) k oddělení funkčních tRNA-like a mRNA-like domén ve struktuře tmRNA (záchrana ribozomu z aberantní mRNA u bakterií)
Funkce RNA • Typy RNA • 1) mRNA • 2) „udržovací“ (housekeeping) RNA • - tRNA, rRNA, snRNA,snoRNA, telomerázová RNA, SRP RNA, RNasaP, RNAsa MRP, tmRNA… • 3) regulační RNA • miRNA (micro), siRNA (short interfering), piRNA (piwi interacting), riboswitch, sRNA 4)„parazitické“RNA (retrotranspozony, viroidy, virusoidy, RNA virů) 5) jiné RNA (guide RNA) • Fce. RNA • - Čtení genetického kódu • - Primery pro replikaci DNA • - Genom mnoha virů • - Strukturní (ribozom, SRP…) • - Ribozymy (katalytická fce.) • Riboswitch (metabolite-sensing) • RNA interference (siRNA, miRNA, piRNA…) Červeně: ribozymová aktivita (u snRNA a rRNA jen některé – U2+U6 snRNA, 23S rRNA)
RNA Tie Club (24 členů s kravatou báze nebo AMK) (foto: zleva Crick, Rich, Orgel, Watson) • Adaptorová hypotéza: • informace v DNA není převáděna do polypeptidu přímo • vodíková vazba mezi nt+AMK je nepravděpodobná • existuje tRNA = adaptorová molekula, která se kovalentně váže k AMK a nekovalentně k NK • Mimo klub: 1953 – Zámečník a kol. • Užití cell-free extraktů, radioaktivně značených AMK a ultracentrifugy • Objev rozpustné S (soluble) RNA (což byla tRNA) • Na S-RNA se nejdříve váže AMK a až pak se AMK dostanou do proteinů
Objev rRNA a mRNA (1960) • rRNA (85% RNA v buňce), je v ribozomech = templát?? • NE – uniformní délka + kompozice bází (GC-bohaté) u různých organismů • objev mRNA na základě infekce E. colifágem T4 • fágová RNA má velmi podobnou kompozici bází jako fágová DNA • RNA se neváže s ribozomálními proteiny za tvorby ribozomálních partikulí • pohybuje se napříč povrchem ribozomu • templát, který určuje pořadí AMK v proteinu • variabilita v délce a kompozici bází
Severo Ochoa – 1959 NC za syntézu RNA - polynukleotid fosforyláza (PNPáza) – in vivo u bakterií štěpí RNA na mononukleotidy, ale přidá k nim navíc ještě 1 fosfát – vznikají nukleosiddifosfáty 1. Kodon – Phe(Nierenberg, Matthaei, 1961; NC roku 1968) Užití PNPázy na syntézu poly -U templátu z nukleosiddifosfátů Translace in vitro z poly-U mRNA- produktem byl polyfenylalanin …pak CCC, AAA…(GGG nefungoval)
Genetický kód 4 báze a kód je tripletový = 3 báze kódují 1 aminolyselinu (AMK) = 43 kombinací = 64 kodonů, z toho 61 kóduje AMK, 3 stop kodony; kodon pro methionin je zároveň iniciační Degenerovaný kód = více různých kodonů kóduje stejnou AMK Univerzální = až na výjimky je stejný u všech organismů Nepřekryvný = báze 1,2,3 kódují 1AMK, báze 4,5,6 kódují další, není tam překryv
Modifikované báze Wobbling inosinu v antikodonu
Ribozymy – objev (80.léta, NC 1983) GI intron ve 26S rRNA Tetrahymena thermophila (Thomas Cech) Rnáza P – sestřihnutí 5´konce prekurzoru tRNA u E. coli (Sidney Altman)
Ribozymy Ribozymy obecně často něco štěpí (s výjimkou např. ribozomu, který syntetizuje), štěpení probíhá v jednovláknové oblasti, obvykle je přímo či nepřímo zůčastněna 2´OH skupina, proteiny udržují správnou konformaci ribozymu, pro funkci je dále důležitá vazba kationtů Hammerhead ribozym u viroidů a virusoidů
Riboswitch Centrální dogma MB Genetika - český překlad (2009) Principles of Genetics 5th edition (Snudtad, Simmons, 2009)
RNA svět • Ribozymy a riboswitche – náznak, že původní svět mohl být založen na RNA • DNA – stabilnější (bez 2´OH, T místo U) – přesun odpovědnosti za uložení genetické informace • Proteiny – převzaly většinu katalytických rolí • Chybí důkazy RNA světa (fosílie)