Download

1 / 30
360 likes | 1.24k Views
Bioenergetics and Metabolism: Energy Input Restrains Entropy Metabolic Flux is Highly-Regulated . Bioc 460 Spring 2008 - Lecture 24 (Miesfeld). Life on earth requires energy from the sun. The flux of people through the DC metro is similar to the flux of metabolites in cells.

E N D
Bioenergetics and Metabolism:Energy Input Restrains Entropy Metabolic Flux is Highly-Regulated Bioc 460 Spring 2008 - Lecture 24 (Miesfeld) Life on earth requires energy from the sun The flux of people through the DC metro is similar to the flux of metabolites in cells Entropy wins every time
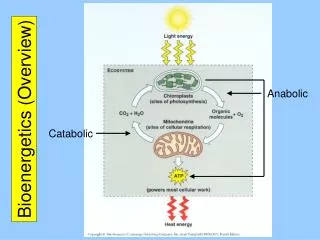
Key Concepts in Bioenergetics and Metabolism • Living organisms must have a constant input of energy in order to maintain a homeostatic condition that is far from equilibrium: an organism at equilibrium with the environment is no longer alive. • Sunlight is the ultimate source of energy for life on earth. Photosynthetic organisms convert sunlight energy into carbohydrates using redox reactions. Carbohydrates are a rich source of redox energy which is converted to phosphoryl transfer energy in the form of ATP. • The Energy Charge (EC) is a measure of [ATP], [ADP], and [AMP] in the cell and reflects the amount of available ATP for metabolic reactions. A low EC simulates flux through catabolic pathways that replenish ATP levels, whereas, a high EC stimulates flux through anabolic pathways to build up carbohydrate and lipid stores.
An organism at equilibrium with the environment is no longer alive Energy is required to keep the level of glucose higher inside the saguaro than outside in the desert. Similarly, energy is required to keep the level of NaCl lower inside the whale than in the sea water. What is the source of energy for the saguaro, what about the whale?
Energy conversion in biological systems is used to perform work • Chemical work in the form of macromolecular biosynthesis of organic molecules. • Osmotic work to maintain a concentration of intracellular salts and organic molecules that is different than the extracellular milieu. • Mechanical work in the form of flagellar rotation or muscle contraction.
Cycling of resources and waste between environment and a living cell provides materials for energy conversion Photosynthetic autotrophs use solar energy to oxidize water and generate chemical energy that us used to convert CO2 into carbohydrate (C6H12O2). Heterotrophs are dependent on photosynthetic autotrophs, or other heterotrophs, to provide the chemical energy needed for life. The byproducts of aerobic respiration are CO2 and water.
Metabolic redox reactions • Both photosynthesis and aerobic respiration interconvert energy using a series of linked oxidation and reduction reactions, also called redox reactions, to transfer electrons (e-) between metabolites. • Oxidation Is Loss of electrons and Reduction Is Gain of electrons (OIL RIG). • Since electrons cannot exist free in solution, they must be transferred between compounds, this transfer process is controlled by enzymes in biological systems.
Redox reactions are catalyzed by enzymes that permit the energy to be captured as work
Aerobic respiration uses redox reactions to interconvert chemical energy available from glucose oxidation into stored energy as ATP Nicotinamide adenine dinucleotide (NAD), flavin adenine dinucleotide (FAD), and quinone (Q) are electron carriers in numerous biochemical reactions.
Thermodynamic principles govern energy conversion in metabolic processes • First Law of Thermodynamics: The total amount of energy in the universe does not change; energy can neither be created or destroyed. Energy conversion in biological systems is never 100%. • Second Law of Thermodynamics: In the absence of energy input, all natural processes in the universe tend toward disorder, the measure of this disorder, or randomness, is called entropy (S). • Gibbs Free Energy (G) and Equilibrium Constant (Keq): In the reaction A-->B, ΔG = ΔGº’+ RT • ln [B]equilibrium [A]equilibrium
First Law of Thermodynamics Energy cannot be created or destroyed – it can only be converted between forms. Antoine Laurent Lavoisier and the guinea pig experiment, 1783. The amount of water collected from melting ice was a measure of metabolic heat produced by the guinea pig.
The energy available for oxidation of a reduced compound in a biological system can be determined precisely using a device called a bomb calorimeter. A bomb calorimeter measures the change in temperature that occurs when a compound is completely oxidized. For example, the amount of stored energy in glucose. C6H12O6 + 6 O2→ 6 CO2 + 6 H2O + heat
The definition of a Calorie • A Calorie (kcal) is defined by the amount of heat energy required to raise 1 kilogram of water from 14.5 ºC to 15.5 ºC at 1 atmospheric pressure. • This can also be expressed in the international unit of measurement the Joule (J) in which 1 Calorie = 1 kcal = 4.184 kJ. • Note that Calorie is sometimes written as calorie in nutrition, but in most cases, it is actually referring to a Calorie (kcal).
Total energy potential of 1 gram of glucose is the same regardless of the metabolic path taken, i.e., 15.7 kJ of energy. We can use this experimental value to calculate metabolic energy available from glucose oxidation. Note that the total amount of energy available does not tell you if glucose oxidation is favorable or not, for that, you need to know the change in Gibbs Free Energy (G).
Second Law of Thermodynamics All natural processes in the Universe tend towards disorder, the measure of which is entropy (S). When S>0, then randomness is increasing. Energy is required to restrain the natural tendency toward disorder, i.e., to keep S from increasing in a system, energy must be provided. An increase in entropy happens when a cell dies and its contents equilibrate with the environment. To stay alive, one must maintain order and prevent the system (cell) from reaching equilibrium with the environment.
The physical state of H2O is an example of how entropy and energy are related The H2O molecules become more and more disordered as heat is added to the system which increases the entropy. Ice melting at room temperature is spontaneous however, this process can be restrained by keeping the ice in a freezer that is kept below 0º C using the energy of electricity.
Gibbs Free Energy is a term relating enthalpy, temperature, and entropy in a way that predicts the spontaneity of a reaction Since the absolute values for enthalpy (H) and entropy (S) cannot be easily determined, the Gibbs Free Energy term (G) is defined as the change between two states: G = H - TS • The value of G for a reaction reveals whether it is favorable or not in the direction written: • If G < 0, the reaction is favorable and exergonic • If G > 0, the reaction is unfavorable and endergonic • If G = 0, the reaction is at equilibrium
The equilibrium constant, Keq, is the ratio of the product and reactant concentrations when the reaction has reached equilibrium At equilibrium, under constant temperature and pressure, the Keq is also a measure of reaction directionality since a Keq > 0 means product formation is favored, whereas, a Keq < 0 means that reactant formation is favored. A + B C + D Keq = [C] [D] [A] [B] Gibbs defined the relationship between G and Keq using a term called the standard free energy change, Gº (kJ/mol), in terms of the gas constant (R) and absolute temperature (T): G = Gº + RT ln Keq
The importance of G in metabolism is that it can be used to predict the direction of a reaction under physiological conditions The Gº’ value (used by biochemists) is experimentally determined by setting up the reaction under standard physiological conditions (pH 7, 55.5 M H2O, 298ºK, 1 atmosphere pressure, and 1 M of each component), in the presence of enzyme, and letting the reaction go to equilibrium. Once the reaction has reached equilibrium, G is 0, and Gº’ can be calculated by measuring [reactant] and [product]: G = Gº’ + RT ln Keq = 0 Gº’ = -RT ln Keq If the Gº’ < 0, the reaction is favorable as written, if Gº’ > 0, the reaction is unfavorable in the direction written.
How do G and Gº’ relate to metabolism? The G of a reaction in a cell is calculated using the Gibbs Free Energy equation and the mass action ratio which is a measure of the actual concentration of reactants and products under physiological conditions. Remember that energy available for work comes from the fact that most chemical reactions inside cells are not at equilibrium (equilibrium is death). G = Gº’ + RT ln [products]actual [reactants]actual How can the G for a reaction in a cell be favorable even though the Gº’ is unfavorable? What can be said about rate of a reaction based on the value of Gº’?
Coupled reactions share metabolic intermediates and combine endergonic and exergonic reactions to yield an overall G that is favorable For example, the product of endergonic reaction (A ↔ B) is the substrate for an exergonic reaction (B ↔ C) to yield an overall favorable reaction that converts reactant A to product C. The ΔG of this coupled reaction is the sum of the ΔG values for each individual reaction and is favorable, thereby, driving the reaction A ↔ C. The same is true if we were using Gº’ values for these reactions. A ↔ B ΔG = +4 kJ/molB ↔ C ΔG = -10 kJ/mol A ↔ C ΔG = -6 kJ/mol
One type of shared reaction is one in which ATP hydrolysis (phosphoryl transfer) is used as a source of energy to drive unfavorable reactions ATP hydrolysis: ΔGº’ = - 30.5 kJ/mol The first step in glycolysis is catalyzed by the enzyme hexokinaseand utilizes phosphoryl transfer from ATP to drive the unfavorable reaction of glucose phosphorylation in a coupled reaction Glucose + Pi ↔ glucose 6-phosphate + H2O ΔGº’ = +13.8 kJ/molATP + H2O ↔ ADP + Pi ΔGº’ = -30.5 kJ/molGlucose + ATP ↔ glucose 6-phosphate + ADP ΔGº’ = -16.7 kJ/mol
Metabolic flux is a term that explains the flow of metabolites through a set of link reactions By combining endergonic reactions with exergonic reactions, some of which can be regulated by controlling enzyme activities, and by altering the concentrations of reactants and products as a result of dietary intake, the flux of metabolites through linked reactions can vary. The two most important considerations for metabolic flux are: 1) the concentration of reactants and products in the cell 2) enzyme activity levels. A ↔ B (G < 0) the rate of B is favored when [A] is high B ↔ C (G = 0) the reaction is sensitive to [B] and [C] C ↔ D (G > 0) the enzyme X is highly regulated A --> D under conditions where [A] is high and enzymes X is active
An example of an ATP-coupled reaction in which both substrate concentration and enzyme activity combine to increase metabolic flux A combination of two reactions in the glycolytic pathway show how conversion of glucose-6-phosphate to fructose-1,6-bisphosphate (BP) occurs in cells even though the Gº’ for each uncoupled reaction is unfavorable (not counting the Gº’ for ATP hydrolysis). Glucose-6-P ↔ Fructose-6-P + ATP ↔ Fructose-1,6-BP + ADP Glucose-6-P ↔ Fructose-6-P Gº’ = +1.7 kJ/mol Fructose -6-P + Pi ↔ Fructose-1,6-BP Gº’ = +16.3 kJ/mol ATP ↔ ADP + Pi Gº’ = -30.5 kJ/mol Glucose-6-P + ATP ↔ Fructose-1,6-BP + ADP Gº’ = -12.5 kJ/mol
The actual concentration of glucose-6-P in the cell under homeostatic conditions is 8.3 x 10-5M, and that of fructose-6-P is 1.4 x 10-5M. Using Gibbs Free Energy equation, and the mass action ratio, the G for the conversion of glucose-6-P to fructose-6-P by the enzyme phosphoglucose isomerase can be calculated. Glucose-6-P ↔ Fructose-6-P G = Gº’ + RT ln [Fructose-6-P]actual [Glucose-6-P]actual G = +1.7 kJ/mol + RT ln 1.4 x 10-5M 8.3 x 10-5M G = -2.9 kJ/mol Phosphoglucose isomerase
Since the phosphofructokinase 1 reaction uses phosphoryl transfer energy available from ATP, it is a highly favorable reaction that uses up fructose-6-P and “pulls” the phosphoglucose isomerase reaction to the right. How would metabolic flux be affected if more glucose-6-P were added?
Metabolic flux is affected by the availability of ATP which is needed as a substrate for numerous coupled reactions in the cell The adenylate system describes the relationship between ATP, ADP, and AMP and can be described as the “energy charge” in the cell. The energy in the two ATP phosphoanhydride bonds ( and ) can be used not only to stimulate catalysis and metabolic flux, but also to induce protein conformational changes and to alter protein-protein interactions sites. Cleavage of the phosphoanhydride bonds in ATP is favored for several reasons: • unstable due to PO4- • entropy increases • increased solvation • release of H+
The Energy Charge of the Cell ATP is continually being hydrolyzed and resynthesized from ADP + Pi. The average person cycles through 50 kg of ATP every day!
Adenylate forms are recycled in the cell by two major biochemical processes; the adenylate kinase reaction and phosphorylating systems Converting AMP to ADP to provide substrate for phosphorylation reactions is the job of adenylate kinase. The primary phosphorylating systems in nature are oxidative phosphorylation (aerobic respiration) and photophosphorylation (photosynthesis). Both of these processes require energy input to drive the ATP synthase reaction. AMP + ATP ↔ 2 ADP (Adenylate kinase; G = ~0 kJ/mol)2 ADP + 2 Pi ↔ ATP + ATP (ATP synthase; G = +61 kJ/mol) AMP + 2 Pi ↔ ATP Where does the energy come from in aerobic respiration and photosynthesis to drive the ATP synthase reaction forward?
The Energy Charge in the cell is maintained between 0.7 and 0.9 as are result of regulated flux through anabolic and catabolic pathways
Relative metabolic flux through catabolic and anabolic pathways responds to changes in the Energy Charge in the cell to maintain homeostasis What do you think the primary mechanism is that regulates metabolic flux through catabolic and anabolic pathways in response to EC?