The Adaptive Immune Response
330 likes | 683 Views
The Adaptive Immune Response. Chapter 16. Note: In this class, antibody and immunoglobulin are synonymous. The Adaptive Response is Slow to Start.

The Adaptive Immune Response
E N D
Presentation Transcript
The Adaptive Immune Response • Chapter 16
Note: In this class, antibody and immunoglobulin are synonymous
The Adaptive Response is Slow to Start • During childhood, billions of T cells and B cells (lymphocytes) are generated, but only a few of these can recognize a given species of pathogen • When these cells recognized the particular pathogen, they begin to rapidly divide that results in a dramatic increase of pathogen-specific cells over several days - termed clonal expansion • A single lymphocyte can divide in about 8 hours (3 divisions/day) • In 5 days, 215 divisions occur, resulting in 32,768 cells • During these first few days of infection, the pathogen can cause substantial pathology to the individual • If the pathology is severe enough, it can cause the death of the individual • Because of this delay, it is critical that the innate immune response immediatelyengage the pathogen and delay its actions long enough for the adaptive response to join the battle
16.1 Strategy of the Adaptive Immune Response • All microbial components are potential antigens for the immune response • Antigens are any molecules that can elicit an immune response • Primary Immune Response • The first exposure to an infectious agent • Slow to start • Often with pathology • Secondary Immune Response • Any additional exposures to the pathogen • Very quick to respond - usually without symptoms
Innate Adaptive Type I Type II 16.1 Strategy of the Adaptive Immune Response • The Two Adaptive Immune Responses • Type I immunity (formerly cell-mediated immunity) • Type II immunity (formerly humoral or antibody immunity) • These responses are often mutually exclusive Immunity
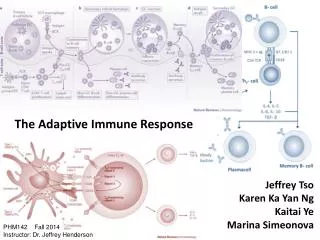
16.1 Strategy of the Adaptive Immune Response • Overview of Humoral (Type II) Immunity • B cells are covered with thousands of copies of a membrane-bound immunoglobulin, termed B cell receptor (BCR) or surface Ig (sIg) • The immunoglobulin is specific for a particular molecule from a particular species of pathogen • If the pathogen enters the body, then this cell will bind to it (or its products) by virtue of its BCR • This results in the activation and maturation of the B cell into a plasma cell, which then begins to secrete soluble versions of its immunoglobulin • These antibodies can circulate in blood and lymph for years • If they bind to the antigen, they target it for destruction
16.1 Strategy of the Adaptive Immune Response • Overview of Cellular (Type I) Immunity • Subsets of T cells • Helper T (Th) cells • Secretes cytokines that mediate local immune responses • Augment activities of the innate response • Are the “generals” of the immune response • Cytotoxic T (Tc) cells • Physically interact with cells harboring a pathogen (such as a virus) and kills the cell • Are the “special forces” of the immune response • Regulatory T (Treg) cells • Suppress inflammation as the immune response wanes
16.2 Anatomy of the Lymphoid System • Lymphatic vessels contain lymph • Secondary lymphoid organs are where immune cells gather to coordinate their activities • Lymph nodes • Spleen • Peyer’s patches (large intestine) • Many others • Primary lymphoid organs are where immune cells develop from stem cells • Bone marrow • Thymus
16.3 The Nature of Antigens • Antigen composition • Protein • Lipids linked to other macromolecules • Carbohydrates linked to other macromolecules • A microbe usually has tens to thousands of antigenic determinants (aka, epitopes) that can be recognized by the immune system
16.4 The Nature of Antibodies • Antibodies are glycoproteins (carbohydrates and polypeptides) • Structure and Properties • Two arms that compose the Fab (fraction antibody) region that bind to antigenic determinants (the “business end”) • Antibodies are divalent • One stem (Fc - fraction crystalizable) that can bind to phagocyte receptors or complement proteins
16.4 The Nature of Antibodies • Each antibody is composed of • Two identical light (L) polypeptide chains • Two identical heavy (H) polypeptide chains • These polypeptides have intrachain and interchain disulfide bonds (covalent) that hold the chains together and maintain the antibody’s 3D shape • There are 5 antibody classes based upon the H chains • IgG • IgM • IgE • IgA • IgD The class of the antibody dictates its biological functions
16.4 The Nature of Antibodies • The variable regions of H and L chains dictate binding to the antigen • The constant regions of H and L chains determine the 3D structure and the biological activities of the antibody
16.4 The Nature of Antibodies • Protective Outcomes of Antigen-Antibody (Ag:Ab) Binding • Antibodies bind to antigens with noncovalent interactions • Early in the adaptive response, antibody affinity is weak, while late in the response it is very strong • Effects of antibody binding to antigen • Neutralization - blocks the biologically-relevant portion of an antigen, rendering it inactive • Immobilization and preventing adherence • Agglutination and precipitation • Opsinization facilitates phagocytosis • Complement system activation • Antibody-dependent cell cytotoxicity (ADCC)
16.4 The Nature of Antibodies • IgG • Most common Ab in serum • High affinity • Several subclasses • Some are efficient complement fixers • Others mediate ADCC • Some cross the placenta and are in breast milk (colostrum) • Immunoglobulin classes • IgM • First secreted antibody • Exists as a pentamer (valency of 10) • Efficient at complement fixation • Primary antibody against LPS • Low affinity, but high avidity • Avidity is the collective strength of interaction
16.4 The Nature of Antibodies • IgD • Usually expressed as BCR • No known functional role • IgE • Only small amounts are produced • Efficient at inducing degranulation of granulocytes • May be protective against helminths • Associated with allergies • Immunoglobulin classes • IgA • Most abundant Ab • Exists as monomers (blood/tissues) and dimers (secreted) • Breast milk • The J chain holds two IgA Ab together to form the dimer and provides protection from digestive enzymes
16.5 Clonal Selection and Expansion of Lymphocytes • Clonal selection theory states that antigens select T and B cells for expansion • The presence of antigen stimulates the naive T and B lymphocytes into mitosis and differentiation that leads to a large pool of antigen-specific effector lymphocytes • As the infection is cleared, antigen becomes limiting and most of the responding lymphocytes die (99% or more) • The remaining lymphocytes serve as memory cells, poised to engage the pathogen should it recur
16.6 B Lymphocytes and the Antibody Response • Antigens recognized by B cells • T-independent (do not require T cell help) • Non-proteinaceous (e.g., LPS) • Cannot class-switch from IgM to others • Do not induce durable immunity • T-dependent (requires T cell help) • Proteins • Class switching to IgG, IgA, IgE • Usually provide long-term immunity
B cell activation to protein antigen • A B cell has thousands of identical BCR molecules protruding from its surface • These BCR have identical specificity (clonotypic) because they have the exact same amino acid sequences • If a protein antigen is bound by these BCR, it is internalized • The internalized antigen is fragmented into peptides of 10-15 amino acids • The peptides are assembled on MHC class II proteins, which are then presented on the B cell surface • A helper T cell that recognizes the peptide engages the B cell to activate it by: • Releasing cytokines (noncognate signals) • Cognate (physical) interactions with the B cell • The B cell begins secreting antibodies (which are soluble versions of the BCR)
16.6 B Lymphocytes and the Antibody Response • Characteristics of the primary B cell response • Class-switching to other isotypes • Affinity maturation by programmed mutation of antibody variable genes • Most mutations result lower or no change in affinity of BCR • Some result in higher affinity BCR and these out compete B cells with lower affinity • Generation of memory B cells • These three events require T cell help
16.6 B Lymphocytes and the Antibody Response • Characteristics of the secondary B cell response • The memory cells, which can number in the tens of thousands, are capable of rapid response to recurrent infections • They do not have to undergo class-switching • They do not have to undergo affinity maturation • They undergo clonal expansion again • This occurs so rapidly, symptoms of illness are rare
16.6 B Lymphocytes and the Antibody Response • The response to T-independent antigens • Reliant upon repetitive motifs of the antigen on the surface of the microbe • Results in cross-linking multiple BCR • This causes B cell activation and secretion of soluble antibody (IgM only)
16.7 T Lymphocytes: Antigen Recognition and Response • General characteristics of T cells • Possess a clonotypic T cell receptor (TCR) • The TCR is virtually identical to one Fab antibody arm • Recognize antigenic peptide fragments of microbial proteins • The peptide fragments must be bound to a major histocompatibility complex (MHC) molecule • MHC class I molecules are for presenting peptide antigens from intracellular microbes (i.e., infected cell) and target those cells for destruction by Tc cells • MHC class II molecules are for presenting peptide antigens from extracellular microbes (i.e., NOT infected cell) and presenting those peptides to Th cells, which secrete cytokines that mediate the local immune response
16.7 T Lymphocytes: Antigen Recognition and Response • The MHC I pathway targets a cell for destruction • Intracellular pathogens use cellular ribosomes to synthesize their proteins • Some of these proteins are fragmented and inserted into the lumen of the ER • MHC I proteins bind to the peptides and then are displayed on the cell’s surface • CD8+ Tc cells recognize these microbial peptides and kill the cell • Puncturing holes in the membrane with perforin • Inducing a death signal that causes DNA fragmentation
The MHC II pathway is for activating Th cells that secrete cytokines to mediate immune responses during infections • Only professional antigen presenting cells (APC), such as macrophages, dendritic cells, and B cells, participate in this pathway • They captureextracellular antigens and internalize them • B cells use their BCR • Macrophages use phagocytosis • Dendritic cells use pinocytosis • The antigens are fragmented into peptides phagolysosomes that possess MHC II proteins, which bind to the peptides • The MHC II:peptide complex is displayed on the cell surface • CD4+ Th cells recognize the peptide and respond by making cytokines • The APC is not killed
16.7 T Lymphocytes: Antigen Recognition and Response • Subsets of CD4+ Th cells • Th1 cells secrete inflammatory cytokines • Th2 cells secrete cytokines that augment B cell responses • Treg (regulatory) cells secrete anti-inflammatory cytokines • The responding subset can determine susceptibility or resistance to a pathogen