Download

1 / 1
10 likes | 190 Views
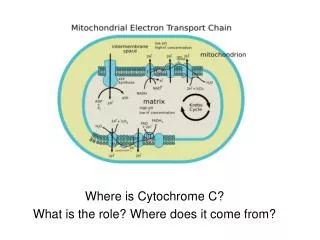
Isolation and Characterization of Cytochrome C-556 from Halothiobacillus neapolitanus. PRANATHI CHEEMAKURTHI and NEWTON P.HILLIARD Department of Physical Sciences, Eastern New Mexico University, Portales, NM-88130. Methodology: Cultivation and Harvesting of Bacterial cells:

E N D
Isolation and Characterization of Cytochrome C-556 from Halothiobacillus neapolitanus.PRANATHI CHEEMAKURTHI and NEWTON P.HILLIARDDepartment of Physical Sciences, Eastern New Mexico University, Portales, NM-88130. Methodology: Cultivation and Harvesting of Bacterial cells: Halothiobacillus neapolitanus ATCC 23641 was grown in large scale in a 14L vessel, New Brunswick Scientific Bio Flo 110, with the thiosulfate as the energy source. The conditions maintained in the vessel are aeration with 30LPM of 5% CO2, temperature 30˚C, rotor speed 180rpm, and a continuous supply of medium with 0.4 dilution rates. The pH of the vessel is maintained at 6.95±1 using sodium carbonate (8%w/v). The cells were collected for every 24 hrs in a 50L carboy and stored at 4˚C. These harvested cells were concentrated using Millipore Corporation Pellicon2 tangential flow high performance ultra filtration unit (HPUF) with a 0.5 m2 0.22 micron GVPP membrane (50L to 2L). The resultant cells were centrifuged by using Beckman model J2-21 at 19,000 rpm for 30 minutes, and obtained a pellet of cells which was stored at -20˚C. Lysis of cells and protein extraction: The pellets stored at -20˚C were thawed to room temperature and were lysed by French Pressure Cell Press at 20,000 psi. Repeating the procedure for three times results in lysis of all cells. These lysed cells were centrifuged at 20,000 rpm for one hour. The supernatant liquid contains the soluble protein and this is used for the further procedure. Ion exchange chromatography for isolation of proteins:The soluble protein extract from the lysed cells was poured on to a 5×17cm diethyl-amino ethyl (DEAE) SephaCelTM column (equilibrated with 20mM phosphate buffer, pH 7). The unbound proteins were eluted out of the column. These eluted proteins were again poured on to a 5×8 cm Blue-Agarose column equilibrated with phosphate buffer. Again the unbound proteins were collected which eluted with the buffer. This pass was then taken in to dialysis tubing and concentrated using PEG 20,000 for 24 hrs. This concentrated fraction was then dialyzed against 20mM MES at pH 6. Then it was poured on to a 5×11 cm DEAE-Sepharose TM column equilibrated with 20mM MES at pH 6. The unbound matter is eluted with the buffer. The bound matter was eluted using, 0.2 M salt gradient with MES which was then equilibrated with 20mM TRIS at pH 8. Then it was applied on to a column of 2×10 cm DEAE-Sepharose. Three fractions were eluted with 0.4 M salt gradient, the first of these fractions contained cytochromeC-556. Characterization of cytochromeC-556: UV/Visible spectroscopy: Absorbance spectra in the region of 280-700 nm were taken for both the oxidized and reduced forms of cytochrome C-556. Reduction of the protein was done with sodium dithionite. Molecular weight determination: SDS-PAGE, polyarylamide gel electrophoresis was used to determine the molecular weight. 10% denaturing polyacrylamide gel was prepared by standard method. The gel was run according to the procedure described by Laemmni (1970) with the purified sample and sigma molecular weight marker loaded on it. The gel was then stained with the Coomasie blue and the band was compared with the sigma marker to determine the molecular weight. ABSTRACT: Being a chemolithoautotropic bacteria, Halothiobacillus neapolitanus, will obtain its energy needed for growth and biosynthetic reactions by the oxidation of sulfur. This sulfur oxidation has economic and environmental importance as it can be used for effective removal of reduced sulfur compounds in various microbial leaching processes. This sulfur oxidation pathway contains many enzymes, and the knowledge about this is incomplete. The present purpose is to isolate and characterize the cytochrome C-556 in the sulfur oxidation pathway of H. neapolitanus by using Ion exchange column chromatography. SDS-PAGE results indicate a single size peptide with an apparent molecular mass of 55kDa. Electronic absorption spectra of the reduced form shows room temperature splitting of α-bands with maxima at 548 and 556nm indicating the possibility of multiple haem environments. Additional maxima occur at 522 and 413nm. Table 2. Diagrammatic flow chart of Isolation and purification of cytochrome C-556. • Results: • UV/Visible spectra of the reduced sample showed room temperature splitting of α-bands with maxima at 548 and 556 nm with the additional maxima at 521 and 413 nm. • SDS-PAGE results showed that the cytochrome has a single size of subunit with apparent molecular weight of 55,000 Daltons. Conclusion: From the spectral and SDS-PAGE results it was concluded that the cytochrome C-556 was a single size peptide of high molecular weight with an apparent molecular weight of 55,000 Daltons and with multiple haem environment, inferred by the room temperature splitting of spectra Acknowledgement: This publication was made possible by NIH Grant Number RR-16480 from the BRIN program of the National Center for Research Resources. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of NIH. I would also like to thank Dr. Newton P. Hilliard for his guidance and support. Figure 1 1-Dimensional SDS-PAGE showing the apparent molecular weight of protein. Introduction: Thiobacilli are a group of bacteria that oxidize the sulfur compounds. This oxidation of sulfur compounds catalyzed by Thiobacilli has both negative and positive influences on the environment and economy applications. The applications of these bacteria include desulfurization of gases, waste water treatment, metal leaching and coal desulfurization. Adverse effects are due to the sulfuric acid produced by the bacteria and these are the principle causative agents for acid mine drainage, the major source of ground water pollution in many regions of the United States. Because of their applications these bacteria are industrially important. For the large scale production of these bacteria for industrial uses, a complete knowledge of the sulfur oxidation pathway and the stoichiometry of the oxidation reactions are necessary. But this knowledge of sulfur oxidation path way is incomplete in many species of thiobacilli. The present research, isolation and characterization of the cytochrome C-556 from Halothiobacillus neapolitanus is a part of group research to complete the oxidation pathway in H.neapolitanus. The advantages of choosing these bacteria are the possibility of producing cells in adequate quantities, easy genetic analysis and acid tolerance of bacteria. 205 KDa 116 KDa 97 KDa 84 KDa 66 KDa 55 KDa 45 KDa 36 KDa • Reference: • Kunen, J.G. and Beudeker, R.F. 1982. Microbiology of Thiobacilli and other sulfur-oxidizing autotrophs, mixotrophs, and heterotrophs. Philos Trans R Soc Lond B Biol Sci. 298(1093):473-497 • Roy, A.B. and Trudinger, P.A. 1970. In the biochemistry of inorganic compounds of Sulfur, University press, Cambridge, pp. 1-40. • Schiff, J.A. 1979. Pathways of assimilatory sulfate reduction in plants and microorganisms. Ciba Found. Symp. 72.49-69. • Le Faou, A., Rajagopal, B.S., Daniels, L., and Fauque, G. 1990. Thiosulfate, polythionates and elemental sulfur assimilation and reduction in the bacterial world. FEMS Microbiol. Rev. 6(4):351-381 • Nordstrom, D.K., Alpers, C.N. and Ptacek, C.J. 2000. Negative pH and extremely acidic mine waters from iron mountain, California. Env. Sci. and Technol. 34(2):254-258. • Milde, K., Sand, W., Wolff, W., and Bock, E 1983. Thiobacilli of the corroded walls of the hamburg sewer system. J. Gen. Microbiol. 129:1327-1333. • Colmer, A.R. and Hinkle, M.E. 1947. The role of microorganisms in acid mine drainage. Science 106:253-256. • Martin, E.J. and Hill, R.D. 1968. In Second Symposium on coal mine drainage research, Mellon Institute, Pittsburgh, pp.46-63. • Brierley, C.L. 1982. Bacterial Leaching. Crit. Rev. Microbiol. 6:207-262. Figure 2 Absorbance spectrum of oxidized cytochrome C-556. Figure 3 Absorbance spectrum of reduced cytochrome C-556. Table 1. Sulfur Oxidation Pathways: