Download
1 / 29
300 likes | 747 Views
Cells must either reproduce or they die. Cells that can not reproduce and are destined to die are terminal cells (red blood, nerve cells, muscles cells etc.). The "life of a cell" is termed the cell cycle as there are distinct phases. They are G1, S, G2, M.

E N D
Cells must either reproduce or they die. Cells that can not reproduce and are destined to die are terminal cells (red blood, nerve cells, muscles cells etc.). The "life of a cell" is termed the cell cycle as there are distinct phases. They are G1, S, G2, M
Once a cell comes into existence it is in the G1 or Gap 1. During this time organelles are reproducing, protein synthesis is occurring for growth and differentiation. Because, transcription is occurring, the DNA is uncoiled. This phase is the most variable, ranging from almost nothing to years. The average may be 10 hrs.
S or synthesis is the time that DNA is being replicated. There is some organelle replication as well. This phase ensures that each emerging daughter cell will have the same genetic content as the mother cell.
G2 or Gap 2 is the time from the end of S until the onset of mitosis. During this time, the cell is preparing for mitosis. Proteins are being made and organized for mitosis. A high increase in tubulin for microtubules for spindle fibers.
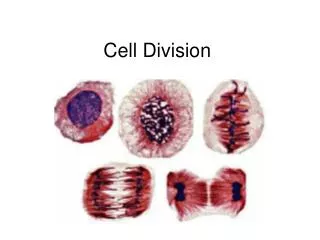
Mitosis or cell division, is the period of time that the nucleus and cytoplasm replicates and divides to produce two genetically identical daughter cells. The phases are triggered by the accumulation of control substances called cyclins.
This graph represents the amount of DNA found in the cell during the cell cycle A –G1 B- S C- G2 D- Mitosis
Internal control of the cell cycle The control of the cell cycle is an accumulation “signal molecules”. These signal molecules must be phosphorylated in order to work. This is a simple model of how this could occur.
Kinases are proteins (enzymes) that phosphorylate these chemical signals or enzymes that trigger the cell cycle phases. This kinase represents the inactive form. This kinase can take two active forms, the S-form or the M-form and phosphorylate different chemical signals. OR
It turns out that cell cycle kinases must be activated by molecules called cyclins. This kinase represents the inactive form with no cyclin attached. The kinase can be activated by molecules called cyclins. This represents two different cyclins. The kinases are called cyclin-dependent-kinases because in order to phos-phorylate the chemical signal, cyclin must be present. This represents what happens when the cyclins are present.
As the cell goes through the cell cycle, different cyclins are made to activate the various cell-dependent-kinases. Once the kinase is activated, the cyclin is destroyed which deactivates the kinase. Kinases are not destroyed, they are only activated or deactivated.
The cell cycle begins. The cell has a certain amount of cyclin-dependent kinases. The cell begins to make the S cyclin.
The CDK complex phosphorylates the S-signal which will initiate the S-phase to start once it gets to a critical level.
Once the S-signal is phosphorylated, it leaves. This causes the S-cyclin to be destroyed and the kinase to return to the inactive state. When there is enough S-signal, then the S-phase will begin.
Now the Cdk is inactive, and the cell begins to make the M-cyclin.
The CDK complex phosphorylates the M-signal which will initiate the M-phase to start once it gets to a critical level. This complex is called the maturation-promoting-factor (MPF).
Once the M-signal is phosphorylated, it leaves. This causes the M-cyclin to be destroyed and the kinase to return to the inactive state. When there is enough M-signal, then the M-phase will begin.
Various cyclins are made and destroyed throughout the cell cycle whereas the level of cell division kinases remain constant. Kinases however are activated by various cyclins and the activity are mirrored by the rise and fall of cyclins.
Fluctuations in concentration of cyclins allow for cell cycle checkpoints. The three major check points are G1/S , G2/M and Spindle checkpoints. These checkpoints have build-in-stop signals that hold the cell cycle at the checkpoint until overridden by go-ahead signals. This is a textbook’s diagram of how cyclins and kinases in the cell cycle work.
Three major checkpoints • G1/S (R point) checkpoint is the primary determining factor for cell division to take place. Growth factors are affecting the cell cycle, and cells are growing. Once the R point is passed the DNA is going to be replicated. If a cell receives a go-ahead signal at this check-point, it will complete the cell cycle and divide. However, if the cell does not receive the go-ahead signal in G1, the switches to a nondividing state called G0.
2. This checkpoint represents the commitment for starting the process of mitosis. This checkpoint also ensures that the DNA has been replicated correctly. If the DNA has been damaged, then the cell does not continue to mitosis. Once the Cdk and cyclin combine, it is called “mitosis promoting factor” or MPF. 3. The M/ spindle check point ensures that all the chromosomes are attached to the spindle in preparation of mitosis. The separation of the chromatids are irreversible. Once chromatids are replicated they are held together by a protein substance called cohesion protein. Another protein called seperase will destroy this protein. Seperase is inhibits or unable to destroy cohesion because of third protein called securin. So in effect the APC (anaphase promoting complex) activates securin, which actives an enzyme seperase to destroy cohesion. In many cells this occurs
during anaphase, however in vertebrates, all of the cohesion is removed during chromatid condensation except the cohesion at the centromeres. Once the cohesion is completely removed, then the tension of the microtubules cause the separation of the chromatids. APC also destroys cyclins in order to drive the cell out of mitosis.
Often the G1 check point or "restriction point" in mammalian cells seems to be the most important one. If a cell receives a go-ahead signal at this check-point, it will complete the cell cycle and divide. However, if the cell does not receive the go-ahead signal in G1, the switches to a nondividing state called G0.
How CDKs actually work is not well understood but the CDKs seem to activate other proteins and enzymes that affect particular steps in the cycle. There has been several discoveries-
External Signals-This include certain chemical and physical factors that affect cell division. Mammalian cells need certain nutrients and regulatory proteins or growth factors are needed for cell division. For example, when the skin has been damage (wound), platelets release a substance called platelet-derived growth factor (PDGF). This growth factor stimulate fibroblast cells to start to reproduce and make scar tissue.
External signals can effect how cells grow in culture. Density-dependent inhibition- cells in culture stop dividing when they become crowded forming a single layer of cells. It seems that when crowded, there is insufficient growth factor produced and nutrients for cell division to continue. Anchorage dependence- mammalian cells need to be attached to substratum like the inside of a culture jar or other tissue in order to reproduce. This phenomenon is linked to a control system attached to the plasma membrane proteins and the cytoskeleton. These phenomenon keep the growth of tissue in check. Cancer cells do not exhibit density-dependent inhibition or anchorage dependence.