1. Background
Direct Amplification and Detection of Enterovirus RNA by Nucleic Acid Sequence Based Amplification (NASBA) A. Heim, J. Schumann, P. Pring-Akerblom Institut für Virologie, Medizinische Hochschule Hannover, 30623 Hannover ahei@virologie.mh-hannover.de.


1. Background
E N D
Presentation Transcript
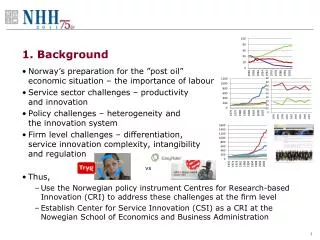
Direct Amplification and Detection of Enterovirus RNA by Nucleic Acid Sequence Based Amplification (NASBA) A. Heim, J. Schumann, P. Pring-Akerblom Institut für Virologie, Medizinische Hochschule Hannover, 30623 Hannover ahei@virologie.mh-hannover.de Figure 2 depicts the positions of the primers and the probe in a multiple alignment of the 5´NTR of various enteroviruses. Moreover, the sensitivity of NASBA was detemined with a panel of laboratory enterovirus strains, and a panel of clinical samples (cerebrospinal fluid and endomyocardial biopsies) which were also screened for enterovirus RNA with a nested RT-PCR [Heim 1997] 3. Results Adjustment of the K+ concentration to 50 mM gave an enhanced sensitivity compared to the frequently recommended K+ concentration of 70 mM (figure 3). Sensitivity was in the range of 10 to 100 molecules positive strand enterovirus (CVB3) RNA with detection of NASBA amplificates by ethidium-bromide stained agarose gel electrophoresis. NASBA was partially selective for positive strand RNA detecting negative strands with an about 100-fold reduced sensitivity (figure 4). 1. Background NASBA is an isothermal nucleic acid amplification reaction (similar to 3SR and TMA) using two specific oligonucleotide primers (one of these contains a T7 promotor sequence), and three enzymes, AMV reverse transcriptase, RNAse H, and T7 RNA polymerase [Compton 1991, Kievits 1991]. In contrast to polymerase chain reaction (PCR), NASBA amplifies RNA directly and does not require an extra reverse transcription reaction. The main reaction product is negative strand RNA. Figure 1 gives an overview of the reaction principle. 2. Material and Methods In order to evaluate NASBA for the detection of genomic enterovirus RNA and to compare its sensitivity with reverse transcription/PCR, the reaction was set up with enterovirus specific primer sequences identical to those proposed by Rotbart [J. Clin. Midrobiol., 1994] for the detection of enterovirus RNA by reverse transcription/ polymerase chain reaction. The upstream primer (QN41, nt. 450-474) was not modified, whereas the downstream primer (QN42, nt. 584-603) had to be modified for NASBA by adding a 5´overhang containing an adapter segment (AGAAGG) and a T7 promotor sequence. NASBA products are mainly negative stranded RNA, and to a minor extent dsDNA (< 10%). These products can be detected by ethidium bromide stained agarose gel electrophoresis or after Northern-blot/slot-blot hybridization with a biotinylated oligonucleotide probe (EV3+) followed by chemoluminescence. Both primers and the probe were synthesized by Eurogentec, Belgium, all other reaction components were provided with the RNamplifire kit (Qiagen, Hilden, Germany). Sensitivity of enterovirus NASBA was determined with invitro transcribed enterovirus RNA (positive strand and negative strand samples). As the KCl concentration of the reaction mix is critical for optimal sensitivity, this had to be adjusted prior to determining the sensitivity. Sensitivity was increased to about 1 to 10 molecules by detection of NASBA amplicons by Northern-blot hybridization (figure 5). A panel of enterovirus strains (prototype strains and clincal isolates) was used to determine the group specifity of enterovirus RNA detection by NASBA (figure 6). Surprisingly, a coxsackievirus B1 strain was not detected, perhaps due to a mutation in the primer binding region. Moreover, a coxsackievirus B5 strain was only amplified faintly. Figure 6. Slot-blot hybridization of NASBA products. Row A: 1 polio1 (Droge), 2 polio2 (Schuster), 3 polio3, 4 CVA7, 5 CVA 10, 6 CVA14, 7 CVA11, 8 CVA21; row B: 1 CVB1 (Schäfer), 2 CVB2 (Ohio 1), 3 CVB4 (Bensehofen), 4 CVB 5 (Kannegiesser), 5 CVB6, 6 echo1 (Farouk), 7 echo2 (Cornelis), 8 echo5 (Noyce); row C: 1 echo6 (Henigst), 2 echo7 (Greon), 3 echo8 (Bryson), 4 echo 9 (Hill), 10 echo11 (Gregory), 11 echo12 (Travis), 12 echo24 (DeCamp) Application of enterovirus NASBA to 30 clinical samples (26 cerebrospinal fluid and 4 endo-myocardial biopsy samples) confirmed a similar sensitivity and specifity as enterovirus RT-PCR. 4 samples were positive by both methods. One sample was positive in the enterovirus NASBA, but not in the nested RT-PCR. This PCR amplicon was identified as CVB6 by nucleic acid sequencing. Another two samples were positive in the enterovirus NASBA, but not in the nested RT-PCR. Allother samples were negative by both methods. 4. Conclusion NASBA is a simple, one step method for the highly sensitive detection of enterovirus RNA, and an excellent alternative method to RT-PCR. 5. References Compton J (1991) Nature,350, 91-92. Heim A et al. (1997) Antiviral Res. 137, 47-56. Kievits T et al. (1991) J. Virol. Methods,35, 273-286. Rotbart HA (1990) J. Clin. Microbiol. 28, 438–442.