The Complement System
The Complement System. Fill free to ask questions during my lectures. In addition, I would also be happy to answer any questions after class. My office is in Room 441 BSB. My email address is boacklrj@musc.edu and my phone number is 792-2550. Robert J. Boackle, Ph.D. .


The Complement System
E N D
Presentation Transcript
The Complement System Fill free to ask questions during my lectures. In addition, I would also be happy to answer any questions after class. My office is in Room 441 BSB. My email address is boacklrj@musc.edu and my phone number is 792-2550. Robert J. Boackle, Ph.D. Please be sure to study the chapter on Complement
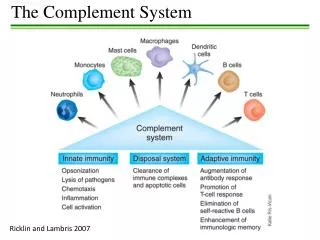
Complement A series of Blood Serum components which exist in a NON-ACTIVE state
The Primary Functions of the Complement System The Primary Functions of the Complement System 1) Quickly Neutralize anything that activates complement by permanently coating the activator 2) Enhance the Phagocytosis of that complement-coated substance 3) Directly damage that complement-coated membrane (e.g., if the activator is a susceptible microbe)
Complement Components are produced Primarily by “HEPATOCYTES” MACROPHAGES and many different cell types also produce complement components
COMPLEMENT Fast Acting (2 min) Potent Tightly Controlled Cascading/Amplifying Sequence
1) FRAGMENTATION 2) CONFORMATIONAL CHANGE 3) ACTIVATION
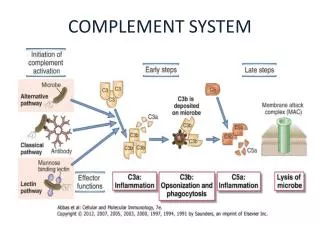
Activation by Immune Complexes The Classical Pathway (Most Powerful Pathway)
Lectin Pathway weakest pathway MBLMASP1 + MASP2 C4b, C3b, iC3b, C3dg, C3d Immune Adherence Enhanced Phagocytosis C3a, C5a Anaphylotoxins Chemotactic Factors Membrane Attack Complex Membrane Damage & Potential for Cell Lysis Classical Pathway Activation by Immune Complexes C1 C4 C2 C3 C5 C6 C7 C8 C9 Membrane damage ) ( (Amplification Loop - Alternative Pathway) C3b, Factor B, Factor D
C3 C5 C6 C7 C8 C9 C3 C5 C6 C7 C8 C9 (Amplification Loop) Cascading - Amplifying Sequence At many steps, especially at C1 and C3 C1 C4 C2 C3 C5 C6 C7 C8 C9 Control at these two steps is important. C4 C2 C3 C5 C6 C7 C8 C9 C4 C2 C3 C5 C6 C7 C8 C9
C1q is a subcomponent of the first complement component C1 C1q Stem region C1q Globular Heads
(Native C1) The first Complement Component, C1 = Macromolecular C1qr2s2 C1q C1r2 and C1s2
C1 C1q Globular Heads bind to the exposed Fc region of IgM, IgG3 or IgG1 Immune Complexes, but not to IgA nor IgG4
= Catalytic Domain Activated C1s Activated C1s C1r C1 Structure, after C1 activation
Movement within the Hinge Region CH2 CH3 CH2 CH3 IgG3 or IgG1
IgG3 or IgG1 ANTIBODIES Movement within the Hinge Region Antigenic determinant CH2 CH3 Foreign (Antigenic) cell/viral surface CH2 CH3 C1q globular heads bind CH2 gamma, but only if the Fab regions have bound antigen and moved so as to expose the CH2 gamma Antigenic determinant
C1 At least two or more IgG antibodies are required but only one IgM (5 Fc regions) At least two adjacent IgG CH2 CH3 CH2 CH3 IgG3 or IgG1 Antigenic Surface
C1q conformationalchanges are induced after binding to adjacent IgG antibodies on the antigenic surface; Followed by C1r2 then C1s2 activation At least two adjacent IgG CH2 CH3 CH2 CH3 IgG3 or IgG1 Antigenic Surface
At least two adjacently deposited IgG antibodies are needed to bind C1, but the more deposited IgG on the antigenic surface the better the interaction with C1 becomes. This requirement for sufficiently deposited levels of antibody insures that complement is not inadvertently activated. However, pathogens that either mutate their immunogenic epitopes or slough their epitopes defeat or circumvent the proper action of the classical complement pathway. IgG3 or IgG1 Antigenic Surface
Active Host Enzymes now on the Antigen C1r2-C1s2 C1q CH 2 Flexibility of C1q FOREIGN SURFACE
C4 Activated C1S 2 FOREIGN SURFACE
C1S 2 C4b Short-lived Active Binding Site FOREIGN SURFACE
C1S 2 C4b Covalent Binding FOREIGN SURFACE Virus Neutralization
C4b that did not bind to the antigen is quickly degraded FACTOR I (protease) C4 Binding Protein C4b If C4b Remains Unbound Note:Bordetellapertussis accumulates serum C4 Binding Protein, resulting in Factor I-mediated inactivation of bound C4b on its surface
C4 C4 C4a C1s C1r2 2 C4b C1q C4b C4b C4b Antigenic Surface
C2 C2 C2b C2a C1s C1r2 C2b 2 C4b2a is a C3 Convertase enzyme complex C1q C2a C2a C2a C4b C4b C4b Antigenic Surface
C4b2a is a C3 convertase enzyme C1s is no longer needed. C1 Inhibitor travels between the now loosened C1q stems and Irreversibly binds and inactivates C1r and C1s C2a C4b ANTIGEN
Native C3 C2 C4b GLY- CYS - GLY- GLU - GLU -THR S C O
Both C4b and C3b are covalently bound to the antigen via the short-lived active binding site. C3b C4b ANTIGEN GLY- CYS - GLY- GLU - GLU -THR H S C O ANTIGEN
Regions on C4b and C3b become “Exposed.” Host Phagocytes attach to these regions on the deposited complement C3a does not bind to antigen, it is released C3b C3a C3b may also attach covalently to C4b on C4b2a C3a C3b C2a C4b As soon as C4b and C3b are Covalently bound to the antigenic surface w w V V Antigenic Surface
PMN or any Phagocyte Immune Adherence & Enhanced Phagocytosis of the complement-coated antigen One of the “major” functions of complement is to enhance Phagocytosis C3b C3b C3b C2a C2a C4b C4b w w w w Antigenic Surface
Each activated complement componenthas at least one inhibitoror inhibitory mechanism Complement Regulation The powerful fast-acting complement system must be controlled at each step or disease ensues due to non-productive depletion of complement.
C1 Inhibitor Release of Ab Release of C1 Dissolution of Immune Complexes C3b C3b C3c C3b C3c C3b C2a C2a iC3b C3d C4b C4b Antigenic Surface Inactivation and removal of C1 occurs as C4b and C3b deposit on the Fab (CH1) or on the antigenic determinants and disrupt the immune complexes, which in turn cause a loosening of the C1qr2s2 complex and allows entrance of C1-Inhibitor. C1-Inhibitor binds and then removes the inactivated C1r and C1s. However, when low levels of deposited IgG are present, the entire C1qr2s2 complex is removed by the action of C1-Inhibitor. Trace amounts of heparin bind to C1q and facilitate the C1-inhibitor mediated inactivation of activated C1r2 and C1s2 and the removal of C1qr2s2 thereby allowing a more efficient maintenance/usage of the (residual) complement components.
Deficiencyof Early Complement Components (C1r, C1s, C4 or C2) Insufficient complement activation on the antigen- No loosening of the Immune Complexes SLE and/or Glomerulonephritis- Inability to clear immune complexes faster than they are forming Possible consequences
C1r C1s Complement Regulation Activated C1, when not bound to immune complexes is rapidly inhibited by C1q C1- Inhibitor Prevents Auto-activation of Native C1 by Activated C1
Activated C1, when not bound to immune complexes is rapidly inhibited by C1r C1s C1- Inhibitor Complement Regulation C1q In the absence of sufficient control by C1-Inhibitor, activated C1r in one (released) activated C1 will activate C1r in other Native C1 (C1 auto-activation) resulting in escalating C1 auto-activation, C4 and C2 consumption and continual depletion of C1-Inhibitor.
C1 Inhibitor DeficiencyGenetic:1. Silent Gene - 25% of Normal C1 INH2. Dysfunctional Gene Product In Both Cases C1 Inhibitor Function is down and C4 levels are lower than normal Acquired: Lymphoproliferative Disorders
Angioneurotic edema (Angioedema) NO Control over unbound activated C1 resulting in total depletion of C4 and C2 and most importantly temporary depletion of C1-inhibitor, resulting in lowered control over Kallikrein and subsequent Bradykinin formation. PlasmaPrekallikrein circulates complexed with high molecular weight kininogen. (Kallikrein activity is controlled by C1 INHIBITOR), Uncontrolled Activated Kallikrein cleaves kininogen to releaseBradykinin.
Complement and Breast Cancer • In Breast Cancer • Elevated Ca++ levels are often observed that may slightly disrupt C1qr2s2, making it more difficult to activate complement on an antibody-coated cancer cell surface • Poor specific antibody responses (to cancer cells) • Elevated levels of complement inhibitory molecules on the surface of the cancer cells • Small levels of complement deposition actually cause surviving cancer cells to become more resistant to apoptosis.
COMPLEMENT (C3b & C4b)Neutralizes Endotoxin Changes the nature of the substance..no longer endotoxin Neutralizes Viruses Virus no longer binds to the target host cell properly
C5a is a “very strong Chemotatic Factor” C5a C5b C5a C5 joins the complex after one or two C3b molecules bind. C5 C3b C2a C3b C4b ANTIGEN
TISSUE INFECTION PMN C5a P M N C5b C3a C3b Concentration Gradient of C5a and C3a
C3a&C5a PMN PMN C5a PMN C3a Complement Coated Antigen
C3a & C5a UP-REGULATION -- Higher Expression of Complement Receptors on PMN PMN
Complement Coated Antigen TISSUE INFECTION PMN P M N ARTHUS REACTION Infected Tissue
C3a and C5a bind to Mast Cells and Basophiles Then Mast Cells and Basophiles release Histamineand Heparin Mast Cell C3a C5a
Histamine Release C3a Mast Cell C5a
Classical Pathway C1 C4 C2 C3 C5 C6 C7 C8 C9 “Membrane damage” LectinPathway Alternative Pathway (Amplification Loop) Membrane Attack Complex (MAC) C5-9
Membrane Attack Complex, C5-C9 C5b C5b Transmembrane Channel C6 C6 C7 C7 C8 Cell Membrane Polymerized C9 C8 enters the membrane, then the polymerized C9 causes the lesion. Then the cell swells.