Download
1 / 1
20 likes | 130 Views
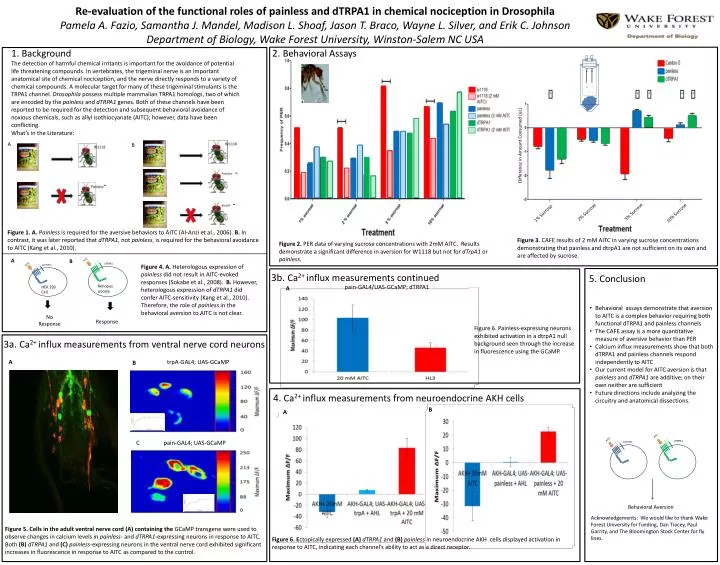
Re-evaluation of the functional roles of painless and dTRPA1 in chemical nociception in Drosophila Pamela A. Fazio, Samantha J. Mandel, Madison L. Shoaf, Jason T. Braco, Wayne L. Silver, and Erik C. Johnson Department of Biology, Wake Forest University, Winston-Salem NC USA. dtrpA1. Painless.

E N D
Re-evaluation of the functional roles of painless and dTRPA1 in chemical nociception in Drosophila Pamela A. Fazio, Samantha J. Mandel, Madison L. Shoaf, Jason T. Braco, Wayne L. Silver, and Erik C. JohnsonDepartment of Biology, Wake Forest University, Winston-Salem NC USA dtrpA1 Painless 1. Background The detection of harmful chemical irritants is important for the avoidance of potential life threatening compounds. In vertebrates, the trigeminal nerve is an important anatomical site of chemical nociception, and the nerve directly responds to a variety of chemical compounds. A molecular target for many of these trigeminal stimulants is the TRPA1 channel. Drosophila possess multiple mammalian TRPA1 homologs, two of which are encoded by the painless and dTRPA1 genes. Both of these channels have been reported to be required for the detection and subsequent behavioral avoidance of noxious chemicals, such as allyl isothiocyanate (AITC); however, data have been conflicting. 2. Behavioral Assays B AITC A AITC painless dTRPA1 What’s in the Literature: C pain-GAL4; UAS-GCaMP Figure 1. A. Painless is required for the aversive behaviors to AITC (Al-Anzi et al., 2006). B. In contrast, it was later reported that dTRPA1, not painless, is required for the behavioral avoidance to AITC (Kang et al., 2010). Figure 4. A. Heterologous expression of painless did not result in AITC-evoked responses (Sokabe et al., 2008). B. However, heterologous expression of dTRPA1 did confer AITC-sensitivity (Kang et al., 2010). Therefore, the role of painless in the behavioral aversion to AITC is not clear. 3b. Ca2+ influx measurements continued 4. Ca2+ influx measurements from neuroendocrine AKH cells Figure 2. PER data of varying sucrose concentrations with 2mM AITC. Results demonstrate a significant difference in aversion for W1118 but not for dTrpA1 or painless. A pain-GAL4/UAS-GCaMP; dTRPA1 A A B Figure 6. Painless-expressing neurons exhibited activation in a dtrpA1 null background seen through the increase in fluorescence using the GCaMP. 3a. Ca2+ influx measurements from ventral nerve cord neurons Xenopus oocyte HEK 293 Cell A No Response Response B Figure 6. Ectopically expressed (A) dTRPA1 and (B) painless in neuroendocrine AKH cells displayed activation in response to AITC, indicating each channel’s ability to act as a direct receptor. AITC AITC dTRPA1 painless Difference in Amount Consumed (uL) trpA-GAL4; UAS-GCaMP B 5% Sucrose 2% Sucrose 1% Sucrose 10% Sucrose Behavioral Aversion Figure 3. CAFE results of 2 mM AITC in varying sucrose concentrations demonstrating that painless and dtrpA1 are not sufficient on its own and are affected by sucrose. Figure 5. Cells in the adult ventral nerve cord (A) containing the GCaMP transgene were used to observe changes in calcium levels in painless- and dTRPA1-expressing neurons in response to AITC. Both (B) dTRPA1 and (C) painless-expressing neurons in the ventral nerve cord exhibited significant increases in fluorescence in response to AITC as compared to the control. 5. Conclusion • Behavioral assays demonstrate that aversion to AITC is a complex behavior requiring both functional dTRPA1 and painless channels • The CAFE assay is a more quantitative measure of aversive behavior than PER • Calcium influx measurements show that both dTRPA1 and painless channels respond independently to AITC • Our current model for AITC aversion is that painless and dTRPA1 are additive; on their own neither are sufficient • Future directions include analyzing the circuitry and anatomical dissections. Acknowledgements: We would like to thank Wake Forest University for funding, Dan Tracey, Paul Garrity, and The Bloomington Stock Center for fly lines.