Download
1 / 1
10 likes | 177 Views
Genetic dissection of nociception in Drosophila melanogaster Madison L. Shoaf, Wayne L. Silver, Erik C. Johnson Department of Biology, Wake Forest University, Winston-Salem, NC shoaml5@wfu.edu. Background

E N D
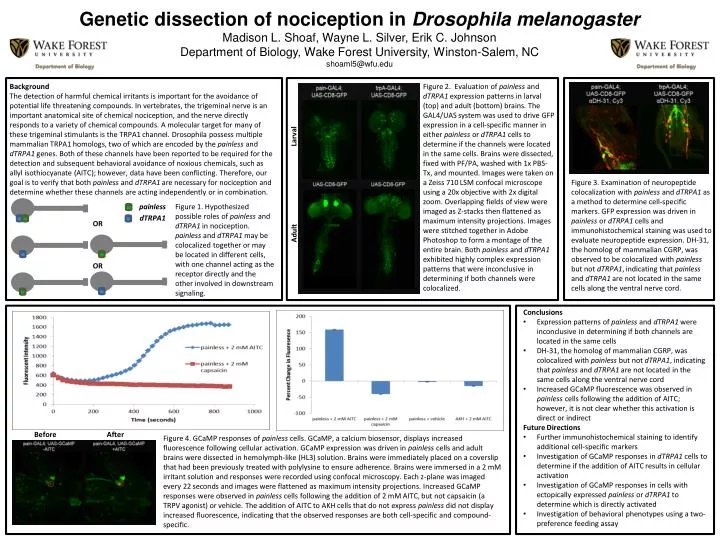
Genetic dissection of nociception in Drosophila melanogaster Madison L. Shoaf, Wayne L. Silver, Erik C. Johnson Department of Biology, Wake Forest University, Winston-Salem, NC shoaml5@wfu.edu Background The detection of harmful chemical irritants is important for the avoidance of potential life threatening compounds. In vertebrates, the trigeminal nerve is an important anatomical site of chemical nociception, and the nerve directly responds to a variety of chemical compounds. A molecular target for many of these trigeminal stimulants is the TRPA1 channel. Drosophila possess multiple mammalian TRPA1 homologs, two of which are encoded by the painlessand dTRPA1 genes. Both of these channels have been reported to be required for the detection and subsequent behavioral avoidance of noxious chemicals, such as allyl isothiocyanate (AITC); however, data have been conflicting. Therefore, our goal is to verify that both painless and dTRPA1 are necessary for nociception and determine whether these channels are acting independently or in combination. Figure 2. Evaluation of painless and dTRPA1 expression patterns in larval (top) and adult (bottom) brains. The GAL4/UAS system was used to drive GFP expression in a cell-specific manner in either painless or dTRPA1 cells to determine if the channels were located in the same cells. Brains were dissected, fixed with PF/PA, washed with 1x PBS-Tx, and mounted. Images were taken on a Zeiss 710 LSM confocal microscope using a 20x objective with 2x digital zoom. Overlapping fields of view were imaged as Z-stacks then flattened as maximum intensity projections. Images were stitched together in Adobe Photoshop to form a montage of the entire brain. Both painless and dTRPA1 exhibited highly complex expression patterns that were inconclusive in determining if both channels were colocalized. Larval Figure 3. Examination of neuropeptide colocalization with painless and dTRPA1as a method to determine cell-specific markers. GFP expression was driven in painless or dTRPA1 cells and immunohistochemical staining was used to evaluate neuropeptide expression. DH-31, the homolog of mammalian CGRP, was observed to be colocalized with painless but not dTRPA1, indicating that painless and dTRPA1 are not located in the same cells along the ventral nerve cord. painless Figure 1. Hypothesized possible roles of painless and dTRPA1 in nociception. painless and dTRPA1 may be colocalized together or may be located in different cells, with one channel acting as the receptor directly and the other involved in downstream signaling. dTRPA1 OR Adult OR • Conclusions • Expression patterns of painless and dTRPA1 were inconclusive in determining if both channels are located in the same cells • DH-31, the homolog of mammalian CGRP, was colocalized with painless but not dTRPA1, indicating that painless and dTRPA1 are not located in the same cells along the ventral nerve cord • Increased GCaMP fluorescence was observed in painless cells following the addition of AITC; however, it is not clear whether this activation is direct or indirect • Future Directions • Further immunohistochemical staining to identify additional cell-specific markers • Investigation of GCaMP responses in dTRPA1 cells to determine if the addition of AITC results in cellular activation • Investigation of GCaMP responses in cells with ectopically expressed painless or dTRPA1 to determine which is directly activated • Investigation of behavioral phenotypes using a two-preference feeding assay Before After Figure 4. GCaMP responses of painless cells. GCaMP, a calcium biosensor, displays increased fluorescence following cellular activation. GCaMP expression was driven in painless cells and adult brains were dissected in hemolymph-like (HL3) solution. Brains were immediately placed on a coverslip that had been previously treated with polylysine to ensure adherence. Brains were immersed in a 2 mM irritant solution and responses were recorded using confocal microscopy. Each z-plane was imaged every 22 seconds and images were flattened as maximum intensity projections. Increased GCaMP responses were observed in painless cells following the addition of 2 mM AITC, but not capsaicin (a TRPV agonist) or vehicle. The addition of AITC to AKH cells that do not express painless did not display increased fluorescence, indicating that the observed responses are both cell-specific and compound-specific.