Download
1 / 1
10 likes | 134 Views
Post DTT. Post DTT. LNRB_orig. HD Domain. LNRB_int. LNR Domain. LNRB_delBC. LNRB_delAB. B. A. Disulfide Connectivity. LNRB_short. A. B. C. 3:6. 2:4. 1:5. A. B. CaCl 2. CaCl 2. A. ZnCl 2. MgCl 2. LNRB_delBC. Mutant LNRB_delBC. LNRB_orig. LNRB_int. Mutant LNRB_orig.

E N D
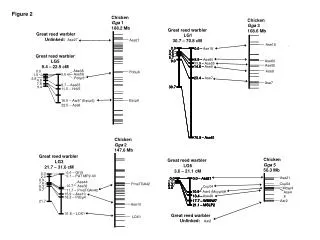
Post DTT Post DTT LNRB_orig HD Domain LNRB_int LNR Domain LNRB_delBC LNRB_delAB B A Disulfide Connectivity LNRB_short A B C 3:6 2:4 1:5 A B CaCl2 CaCl2 A ZnCl2 MgCl2 LNRB_delBC Mutant LNRB_delBC LNRB_orig LNRB_int Mutant LNRB_orig Mutant LNRB_int Characterization of protein folding determinants for LIN-12/Notch-Repeats (LNRs) using Human Notch1 LNR-B as a model system Sharline Madera Advisor: Dr. Didem Vardar-Ulu Wellesley College, Massachusetts NSF REU Chemistry Leadership Group Travel Award Recipient NSF Award CHE-0754512 Abstract Metal Specificity Human Notch1 is a member of a conserved family of heterodimeric type 1 transmembrane receptors that control differentiation in multicellular animals. Notch proteins possess three contiguous LIN-12/Notch-Repeats (LNRs), LNRA, LNRB and LNRC, in their extracellular domain that maintain the receptor in its resting conformation in the absence of ligand. These conserved LNRs display a characteristic disulfide bonding pattern and require Ca2+ for folding. In the receptor, they are separated by two linkers, linker_AB and linker_BC, 10 and 6 amino acids long, respectively. Previously, we had shown that LNRB folding was affected by residues 6-10 of linker_AB and not of linker_BC (1, Figure 5). In this study we investigated the dependence of LNRB folding on the identity of the metal ion as well as the number of conserved disulfide bonds. Our results indicate that LNRB folding is selective for Ca2+ and the linker_AB length requirement and total number of disulfide sulfide bonds needed for effective folding are interdependent. Together these studies represents the initial steps toward defining the minimum requirements for a correctly folding LNR module using LNRB from human Notch1 as a model system. Figure 7. Chromatograms of LNRB_orig and LNRB_int folded under varying metal ion conditions. Panel A: Superposition of chromatograms of LNRB_orig folded in the presence of 10mM CaCl2-red and 10mM MgCl2-black. Panel B: Superposition of chromatograms of LNRB_orig folded in the presence of 1mM CaCl2-green and 1mM ZnCl2-blue. These chromatograms demonstrate the selectivity of LNRB_orig folding for Ca2+ which result in a single folded peak, compared to other potential divalent cations Mg2+ and Zn2+, which yield an array of multiple peaks indicating the lack of one predominant native fold. Figure 4. Sequence Alignment for LNRB Constructs used for the study: Red Residues: Ca 2+ coordinating residues Blue Residues: Mutated cysteines to alanines Orange Residues: Disulfide bonded cysteines linker_BC linker_AB CC CC CC SLNFNDPWKN QRAEGQ LNRA LNRC LNRB Table 1. Summary of HPLC & Mass Spectrometry Results Results Figure 1. Human Notch 1 LNRs and linkers. Introduction Notch Proteins are large Ca2+ binding, transmembrane receptors that control differentiation in multicellular animals. In mammals, there are four Notch homologs: Notch1-4. These proteins function via a highly conserved mechanism referred to as the Notch signaling pathway, which is important for cell-cell communication, involving gene regulation mechanisms that control multiple cell differentiation processes during embryonic and adult life. Deregulation of normal Notch activation has been noted in certain human leukemias, (2) Alagille (3, 4) and CADASIL (5) syndromes, indicating that perturbations of Notch signaling underlie several forms of human diseases (6). Notch proteins exhibit a highly conserved modular architecture (Figure 2), in which distinct repeated structural units are associated with different functional roles in the intact receptor (7). Ligand binding to the N-terminal EGF-repeats activates these proteins by facilitating a proteolytic cleavage by a metalloprotease at site S2, and triggers the gamma-secretase cleavage that permits the translocation of intracellular Notch (ICN) into the nucleus to activate the transcription of target genes (8, 9, 10, 11). The Negative Regulatory Region (NRR) of all Notch receptors has three tandem, independently folding LIN-12/Notch Repeats (LNRs) that wrap around the HD domain containing the regulatory cleavage site S2, and mask the S2 site in the resting receptor (Figure 3) (12-14). Hence the interactions between the LNRs and the HD are critical in stabilizing the NRR and preventing activation prior to ligand binding. Each of the LNRs contains six cysteines with a unique disulfide bonding pattern and coordinate a single Ca2+ (Figure 3), however the minimum requirements that would ensure an LNR to fold independently are not known. This work utilizes different LNRB constructs that all contain the 32 amino acid stretch from cysteine 1 to cysteine 6 in the second LNR of hN1and various numbers of the residues that flank these residues (Table 1), to define the minimum length requirement for hN1 LNRB and to investigate the impact of metal ions and number of disulfide bonds on its autonomous folding. B Figure 5. Chromatograms of LNRB constructs: LNRB_orig- green, LNRB_int- purple, LNRB_delBC- blue.LNRB_delAB- brown, LNRB_short- orangePanel A) Superposition of the chromatograms of the three LNRB constructs that folded into a single predominant peak indicating a single thermodynamically favored correctly folded species. Small neighboring peaks are indicative of misfolded species. Top left inset: Representative chromatogram detailing the elution gradient used and the pressure during the run. Panel B) Superposition of the chromatograms of the two unfolded constructs after dialysis 3. Note no predominant peak is obtained demonstrating no preference for correctly folded species for these two constructs. Top right insets for both panels: Superposition of chromatograms for the same constructs after DTT treatment showing that all peaks of the folded chromatograms collapse to a single peak when reduced and elute later in the gradient (see Table 1). Conclusions In this study we investigated the relative importance of the number of potential disulfide bonds and the divalent ion identity for the independent folding of LNRB in relation to the presence or absence of the linker regions flanking LNRB. Previous folding studies identified constructs LNRB_delBC, LNRB_orig and LNRB_int as autonomously folding constructs. In this study we show two of the modified LNRB constructs where cysteines 1 and 5 are mutated to alanines to eliminate the potential of the first disulfide bond, Mut 1,5C→A LNRB_int and Mut 1,5C→A LNRB_orig, are unable to fold into a single thermodynamically favored folded state. In contrast, Mut 1,5C→A LNRB_delBC retains the ability to fold autonomously. These data show that for the formation of a single thermodynamically favored LNRB species upon in vitro refolding there is a minimum requirement for the total number of stabilizing interactions that participate in the folding process. The relative impact of each of these interactions, which involve linker_AB, the disulfide bonds, and the Ca2+ ion, in ensuring autonomous folding are highly interdependent. Furthermore the attainment of one thermodynamically favored species in the folding of LNRB is dependent and selective for Ca2+ compared to the two other divalent cations abundant in cells, Mg2+ and Zn2+. Future Directions The findings of this study are the initial steps in defining the minimum requirements for an autonomously folding LNR module using LNR_B as a model from human Notch1. They will be followed by studies that include further investigations on the effects of altering the cysteine arrangement of other LNR modules with varying lengths and direct correlation of these findings to hN4LNRA, a wild type LNR module within the human Notch4 receptor and contains only two disulfide bonds. Figure 2. Domain organization of the Notch Receptor. Figure 3. Crystal structure of Human Notch2 NRR (15). Figure 6.Superposition of chromatograms for LNRB_delBC-black and Mutant LNRB_delBC-red. Similar to LNRB_delBC, mutant LNRB_delBC displays a single predominant peak. This finding demonstrates that the mutant LNRB_delBC that contains only two disulfide bonds is capable of folding into a single thermodynamically favored state whose stability is comparable to that of wild type LNRB_delBC. Experimental References • Wildtype hN1LNRB was expressed as inclusion bodies in BL21(DE3)PlysS E.coli. cell line as a fusion protein with a modified form of the trpLE sequence using the pMML vector (kind gift of S. Blacklow, BWH). • The plasmids for the mutant hN1LNRBs were obtained from the corresponding wildtype pMML vector using the QuikChange Site-Directed Mutagenesis protocol (Stratagene) where cysteines 1 and 5 were changed into alanines eliminating the potential for the first disulfide bond. • LNRB was cleaved from the hydrophobic leader sequence by cyanogen bromide cleavage in 70% formic acid and was separated from the leader sequence through precipitation of the leader sequence upon pH increase. • Protein identity for each construct was confirmed by Mass Spectrometry. • Soluble LNRB constructs (~175 M) were folded for two days in a refolding buffer with daily buffer changes. • 100mM NaCl • 20mM Tris pH 8 • 10mM CaCl2/ 10mM MgCl2/ 1mM ZnCl2 • 2.5mM cysteine • 0.5mM cystine • Madera, S. and Vardar-Ulu D. “Characterization of protein folding determinants for LIN-12/Notch-Repeats (LNRs) using Human Notch1 LNR-B as a model system” Poster Presentation, 21st Annual Symposium of the American Protein Society, Protein Society, Boston July 2007. 2. Ellisen, L. W. et al.(1991) Cell. 66:649–661. 3.Li, L., et al. (1997) Nat. Genet. 16: 243–251. 4.Oda, T. et al. (1997) Nat. Genet. 16: 235–242. 5. Joutel, A. et al. (1996) Nature. 383: 707–710. 6. Rand, M. et al. (2000) Molec. and Cell. Biol. 20: 1825-1835. 7. Vardar, D. et al. (2003) Biochemistry. 42: 7061-7067. 8. Sanchez-Irizarry, C. et al. (2004) Molec. and Cell. Biol. 24: 9265-9273. 9. Logeat, F. et al. (1998) Proc. Natl. Acad. Sci. USA. 95: 8108-8112. 10. Brou, C. et al. (2000) Mol. Cell. 2: 207-216. 11. Lawrence, N. (2000) Development. 127: 3185-3195. 12. Aster, J. et al. (1999) Biochemistry. 38: 4736-4742. 13. Weng, A.P. et al. (2004) Science. 306: 269–271. 14. Kopan, R. et al. (2000) Genes Dev.14: 2799-2806. 15. Gordon, W. R. et al. (2007) Nature Structural and Molec. Biol. 14:295-300. • On day 3 the constructs were moved into a dialysis buffer that did not contain any redox reagent (cysteine/cystine). • All constructs from day 3 of dialysis were analyzed on a reverse phase HPLC using a C18 column and 0.25%/min gradient elution under the following experimental conditions: Buffer A: 10% Acetonitrile, 90% H2O, 0.1% TFA Buffer B: 90% Acetonitrile, 10% H2O, 0.1% TFA • A sample of each folded construct was also incubated with 100 mM DTT at room temp for 2 hrs and analyzed by RP-HPLC. Figure 7. Panel A) Superposition of the chromatograms of LNRB_orig-black and Mutant LNRB_orig-red. Panel B) Superposition of the chromatograms of LNRB_int-black and Mutant LNRB_int-red. Unlike the wild type constructs LNRB_orig and LNRB_int, mutant LNRB_orig and mutant LNRB_int do not display a single peak that corresponds to one thermodynamically stable folded state. Instead the chromatograms of the mutant constructs show various peaks indicating the presence of several thermodynamically similar species and not one favored folded state.