Download
1 / 29
330 likes | 910 Views
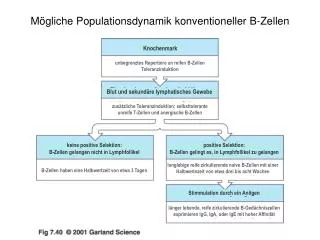
Mögliche Populationsdynamik konventioneller B-Zellen. Knochenmark. unbegrenztes Repertoire an reifen B-Zellen Toleranzinduktion. Blut und sekundäre lymphatisches Gewebe. zusätzliche Toleranzinduktion; selbsttolerante unreife T-Zellen und anergische B-Zellen. keine positive Selektion:

E N D
Mögliche Populationsdynamik konventioneller B-Zellen Knochenmark unbegrenztes Repertoire an reifen B-Zellen Toleranzinduktion Blut und sekundäre lymphatisches Gewebe zusätzliche Toleranzinduktion; selbsttolerante unreife T-Zellen und anergische B-Zellen keine positive Selektion: B-Zellen gelangen nicht in Lymphfollikel positive Selektion: B-Zellen gelingt es, in Lymphfollikel zu gelangen langlebige reife zirkulierende naive B-Zellen mit einer Halbwertszeit von etwa drei bis acht Wochen B-Zellen haben eine Halbwertzeit von etwa 3 Tagen Stimmulation durch ein Anitgen länger lebende, reife zirkulierende B-Gedächniszellen exprimieren IgG, IgA, oder IgE mit hoher Affinität
B-Zellen stammen von einem lymphatischen Vorläufer im Konchenmark ab ein B-Zell-Vorläufer ordnet seine Immunglobulingene um eine unreife B-Zelle, die an ein körpereigenes Zelloberflächenantigen gebunden ist, wird aus dem Repertoire eliminiert eine reife B-Zelle, die an ein Fremdantigen gebunden ist, wird aktiviert aus aktivierten B-Zellen entstehen Plasma- und Gedächniszellen Plasmazelle Gedächniszelle Stromazelle im Knochenmark Sekretion von Antikörpern im Knochenmark und in lymphatischen Geweben sowie Gedächniszellen in lymphatischen Gewebe Entstehung von B-Zellen-Rezeptoren im Knochenmark B-Zellen wandern zu den peripheren lymphatischen Organen negative Selektion im Knochenmark
Die Entwicklung von T-Zellen ein T-Zell-Vorläufer ordnet im Thymus seine T-Zell-Rezeptor-Gene um unreife T-Zellen, die körpereingene MHC-Moleküle erkennen, empfangen Überlebenssignale; andere, die stark mit eigenen Antigenen interagieren, werden aus dem Repertoire herausgenommen reife T-Zellen treffen in peripheren lympatischen Organen auf Fremdantigene und werden aktiviert aktivierte T-Zellen proliferieren und wandern in die Peripherie, um eine Infektion zu verhindern aktiviert tötet positive und negative Selektion im Thymus reife T-Zellen wandern zu den peripheren lymphatischen Organen aktivierte T-Zellen wandern zu Entzündungsstellen T-Zell-Vorläufer entwickeln sich im Knochenmark und wandern zum Thymus
Die frühen Stadien der B-Zell-Entwicklung sind von den Stromazellen des Knochenmarks abhängig Stammzelle / frühe lymphatische Zelle frühe Pro-B-Zelle späte Pro-B-Zelle Prä-B-Zelle unreife B-Zelle Stromazelle im Knochenmark
Die Entwicklung einer Zelle der B-Zell-Line durchläuft mehrere Stadien, die durch Umordnung und Expression der Immunglobulingene charakterisiert sind Stammzelle große Prä-B-Zelle frühe Pro-B-Zelle späte Pro-B-Zelle Keimbahn D-J-Umordnung V-DJ-Umordnung VDJ umgeordnet Gene für die schwere Kette Gene für die leichte Kette Keimbahn Keimbahn Keimbahn Keimbahn Ober- flächen-Ig μ-Kette vorübergehend auf der Oberfläche als Teil eines Prä-B-Zell-Rezeptors; hauptsächlich in der Zelle nicht vorhanden nicht vorhanden nicht vorhanden
Die Entwicklung einer Zelle der B-Zell-Line durchläuft mehrere Stadien, die durch Umordnung und Expression der Immunglobulingene charakterisiert sind kleine Prä-B-Zelle unreife B-Zelle reife B-Zelle Gene für die schwere Kette VDJ umgeordnet VDJ umgeordnet VDJ umgeordnet Gene für die leichte Kette V-J-Umordnung VJ umgeordnet VJ umgeordnet Intrazelluläre μ-Kette Ober- flächen-Ig IgM auf der Zelloberfläche exprimiert IgD und IgM aus alternativ gespleißten Transkripten für die schwere Kette
Die Korrelation von Stadien der B-Zell-Entwicklung mit der Umordnung der Immunglobulingensegmente und der Expression von Zelloberflächenproteinen Stamm-zelle frühe Pro-B-Zelle späte Pro-B-Zelle große B-Zelle kleine B-Zelle unreife B-Zelle Umordnung
Der Zelluläre Aufbei des menschlichen Thymus corticale Epithelzelle Kapsel Trabekel Thymocyt (aus dem Knochen-mark) sub-kapsuläres Epithel Cortex medulläre Epithelzelle Cortico-medulläre Grenze dendritische Zelle (aus dem Knochenmark) Medulla Hassall-Körperchen Makrophage (aus dem Knochenmark)
Die epithelialen Zellen des Thymus bilden ein Netzwerk, das die sich entwickelnden Thymocyten umgibt
Der Thymus ist von entscheidender Bedeutung für das Heranreifen von T-Zellen aus Zellen, die aus dem Knochenmark stammen Stammzellen aus dem Knochenmark Lymphocytendefekt Thymusdefekt Rudimentärer Thymus Thymus-transplantat transplantierte Zellen besiedeln den normalen Thymus normale Zelle besiedeln den transplantierten Thymus Analyse von Milzzellen Analyse von Milzzellen Zellzahl Zellzahl vor dem Trans-plantat vor dem Trans-plantat nach dem Trans-plantat nach dem Trans-plantat Nicht- T-Zellen T-Zellen Nicht- T-Zellen T-Zellen
Im Cortex des Thymus werden sich entwickelnde T-Zellen, die eine Apoptose durchlaufen, von Makrophagen aufgenommen
Aufgrund von Änderungen der Zelloberflächenmoleküle kann man Thymocytenpopulationen verschiedener Reifungsstadien unterscheiden „doppelt negative“Thymocyten große aktive „doppelt positive“Thymocyten Apoptose kleine ruhende „doppelt positive“Thymocyten kleine ruhende „einfach positive“Thymocyten Ausschleusung in die Peripherie
Die Korrelation von Entwicklungsstadien der :β– T-Zellen mit der Umordnung der T-Zell-Rezeptor-Gene und der Expression von Zelloberflächenproteinen einfach positiv doppelt negativ doppelt positiv Umordnung
Die Korrelation von Entwicklungsstadien der :β– T-Zellen mit der Umordnung der T-Zell-Rezeptor-Gene und der Expression von Zelloberflächenproteinen einfach positiv doppelt negativ doppelt positiv einfach positiv Funktion Signalgebung Adhäsions-moleküls IL-2-Rezeptor Signalgebung Corezeptor CD4 oder CD8 unbekannt
In verschiedenen Bereichen des Thymus befinden sich Thymocyten unterschiedlicher Entwicklungsstadien subkapsulärer Bereich unreife doppelt negative CD3¯4¯8¯-Thymocyten Corticale Epithelzelle Cortex unreife doppelt positive CD3+4+8+-Thymocyten Corticomedulläre Grenze Dendritische Zelle reife CD4+8¯ und CD8 +4¯ Medulläre Epithelzelle Thymocyten Medulla Venole Makrophage
Die Bindung an körpereigenen Moleküle im Knochenmark kann zur Eliminierung oder Inaktivierung unreifer B-Zellen führen Unreife B-Zelle (Knochenmark) multivalentes körpereingenes Molekühl lösliches körpereigenes Molekühl nichtquervernetzen-des körpereigenes Molekül mit niedriger Affinität keine Reaktion gegen körpereigene Determinanten klonale Deletion oder Rezeptor-Editing Wanderung zur Peripherie Wanderung zur Peripherie Wanderung zur Peripherie Reife B-Zelle (klonal ignorant) Apoptose anergische B-Zelle Reife B-Zelle
Untersuchungen der positiven Selektion mithilfe von Knochenmarkchimären von Mäusen Übertragung von Knochenmark bestrahlter Empfänger des MHC-Types a bestrahlter Empfänger des MHC-Types b bestrahlt bestrahlt Messung der Reaktion immunisierter F1-T-Zellen auf das Antigen, das von APCs des MHC-Types a und b präsentiert wird Reaktion der T-Zellen Reaktion der T-Zellen immune T-Zellen reagieren auf ein Antigen, das von APCs des MHC-Types a präsentiert wird immune T-Zellen reagieren auf ein Antigen, das von APCs des MHC-Types b präsentiert wird
Zusammenfassung der T-Zell-Reaktionen auf eine Immunisierung von Knochenmarkchimären der Maus Emfpänger sekundäre T-Zell-Reaktion auf das Antigen, das in vitro von APCs des folgenden Typs präsentiert wird Knochenmarks-spender Mäuse mit APCs des Typs: MHCa APC MHCb APC ja nein ja nein nein nein ja nein
Die Epithelzellen des Thymuscortex führen eine positive Selektion herbei normale Expression von MHC-Klasse-II-Molekülen Mutante ohne MHC-Klasse-II-Molekülen CD8- und CD4-Zellen reifen heran nur CD8-Zellen reifen heran Mutante, deren MHC-Klasse-II-Transgen nur im Thymusepithel exprimiert wird Mutante, in der ein MHC-Klasse-II-Transgen exprimiert wird, das nicht mit CD4 interagieren kann CD8- und CD4-Zellen reifen heran nur CD8-Zellen reifen heran
T-Zellen, die auf körpereigene Antigene ansprechen, werden im Thymus eliminiert Transgen Thymus normaler Thymus Thymus und spezifisches Peptid Weit verbreitete Apoptose, zahlreiche apoptische Zellen Ein paar versprengte apoptotische Zellen
Zellen aus dem Knochenmark lösen im Thymus eine negative Selektion aus Knochenmarktransplantat von einer MHC axbF1-Maus in einem MHCa-Emfänger Knochenmark Hauttransplantat von einer MHC b-Maus auf eine (MHCaxb→MHCa) Knochenmarkchimäre Hauttransplantat (MHCaxb→MHCa)-chimäre Maus toleriert ein MHCb-Hauttransplantat Knochenmarkchimäre
Bei der positiven Selektion muss die Spezialität oder Affinität eine andere sein als bei der negativen Selektion Opsitive und negative Selektion haben unterschiedliche Spezialität oder Avidität Opsitive und negative Selektion haben dieselbe Spezialität oder Avidität unreife Thymocyten positive Selektion negative Selektion Reife periohere T-Zellen
Die Unterschiede zwischen negativer und positiver Selektion könnten auf Unterschiede in der Aggregation von T-Zell-Rezeptoren bei der Ligandenbindung zurückzuführen sein reife T-Zelle Sich entwickekelnder Thymocyt Aktivierung der T-Zelle duch ein agonistisch wirkendes Peptid doppelt positiver Thymocyt bindet an agonistisch wirkendes Peptid auf einer Epithelzelle des Thymus Thymocyt stirbt duch Apoptose (negative Selektion) Erkennung eines agonistischen Peptids Antigenpräsentierende Zelle Epithelzelle des Thymis T-Zelle wird durch antagonistisches Peptid gehemmt Doppelt positiver Thymocyt bindet an agonistisch wirkendes Peptid auf einer Epithelzelle des Thymus Thymocyt reift zur einfach positiven CD8-T-Zelle heran Erkennung eines anta-gonistischen Peptids T-Zelle ignoriert Peptid, das weder agonistisch noch antagonistisch wirkt Thymocyt bindet nicht an Peptid, das weder agonistisch nich antagonistisch wirkt Thymoct stirbt durch Apoptose (Tod duch Vernachlässigung) Kein Erkennen (irrelevantes Peptid)
Vergleich der Eigenschaften von B-1-Zellen und konventionellen B-Zellen (B-2-Zellen) Eigenschaft B-1-Zellen konventionelle B-2-Zellen zum ersten Mal produziert Fetus nach der Geburt N-Bereiche in VDJ-Verbindungen wenig zahlreiche Repertoire des V-Bereichs eingeschränkt vielfältig primäre Lokalisation Körperhöhlen (peritonela, pleural) Sekundär lymphatische Organe Art der Ernährung Ersetzt aus dem Knochen-mark selbsterneuernd Spontane Immunglobulinproduktion stark schwach sezernierte Isothypen IgM >> IgG IgG > IgM Reaktion auf Kohlenhydratantigen ja unter Umständen Reaktion auf Proteinantigenen unter Umständen ja Hilfe von T-Zellen erforderlich nein ja Somatische Hypermutation niedrig bis überhaupt nicht stark Gedächtnisentwicklung wenig bis überhaupt nicht ja
Mögliche Populationsdynamik konventioneller B-Zellen Knochenmark unbegrenztes Repertoire an reifen B-Zellen Toleranzinduktion Blut und sekundäre lymphatisches Gewebe zusätzliche Toleranzinduktion; selbsttolerante unreife T-Zellen und anergische B-Zellen keine positive Selektion: B-Zellen gelangen nicht in Lymphfollikel positive Selektion: B-Zellen gelingt es, in Lymphfollikel zu gelangen langlebige reife zirkulierende naive B-Zellen mit einer Halbwertszeit von etwa drei bis acht Wochen B-Zellen haben eine Halbwertzeit von etwa 3 Tagen Stimmulation durch ein Anitgen länger lebende, reife zirkulierende B-Gedächniszellen exprimieren IgG, IgA, oder IgE mit hoher Affinität
Eine Zusammenfassung der Entwicklung menschlicher konventioneller B-Zellen B-Zellen Gene für die schwere Kette Gene für die leichte Kette intra-zelluläre Proteine Marken-protein auf der Oberfläche Stamm-zelle Keimbahn Keimbahn CD34 CD35 RAG-1 RAG-2 TdT λ5, V-Prä-B CD34 CD35 MHC-Klasse II CD10, CD19 CD38 frühe Pro-B-Zelle D-J umgeordnet Keimbahn antigenunabhängig CD45R MHC-Klasse II CD10, CD19, CD38, CD20, CD40 TdT λ5, V-Prä-B späte Pro-B-Zelle V-DJ umgeordnet Knochenmark Keimbahn CD45R MHC-Klasse II Prä-B-R CD CD19, CD38, CD20, CD40 Prä-B-Rezeptor λ5, V-Prä-B große Prä-B-Zelle VDJ umgeordnet Keimbahn CD45R MHC-Klasse II CD19, CD38, CD20, CD40 μ im Cytoplasma kleine Prä-B-Zelle VDJ umgeordnet μ RAG-1 RAG-2 V-J umgeordnet
Eine Zusammenfassung der Entwicklung menschlicher konventioneller B-Zellen intra-zelluläre Proteine Gene für die leichte Kette B-Zellen Gene für die schwere Kette Marken-protein auf der Oberfläche Knochenmark CD45R MHC-Klasse-II IgM, CD19, CD20, CD40 VGJ umgeordnet; schwere μ-Kette in Membranform gebildet VJ umge-ordnet unreife B-Zelle Zu VGJ umgeordnet; μ-Kette in Membranform gebildet; durch alternatives Spleißen entehen μ- und -mlRNA CD45R MHC-Klasse-II IgM, IgD CD19, CD20, CD21, CD40 reife, naive B-Zelle VJ umge-ordnet antigenunabhängig CD45R MHC-Klasse-II CD19, CD20, CD21, CD40 VGJ umgeordnet; alternatives Spleißen führt zur Sekretion von μ-Ketten VJ umge-ordnet Lympho-blast Peripherie VJ umge-ordnet; somatische Hyper-mutation CD45R MHC-Klasse-II IgG, IgA CD19, CD20, CD21, CD40 Isotypenwechsel zu C, C oder Cμ; somatische Hypermutation B-Gedächt-niszelle Isotypenwechsel und alternatives Spleißen führt zur Sekretion von -, -oder μ-Ketten Plasmazell-Antigen- 1 CD38 VJ umge-ordnet Plasma-zelle Letztlich differentziert zu
Eine Zusammenfassung der Entwicklung menschlicher :β-T-Zellen Marken-protein auf der Oberfläche Umordnungen des -Ketten-Gens Umordnungen des β-Ketten-Gens T-Zellen intra-zelluläre Proteine Knochenmark Stamm-zelle Keimbahn Keimbahn CD34? RAG-1 RAG-2 TdT Lck ZAP-70 früher doppelt negativer Thymocyt CD2 HSA CD44hi D-J umgeordnet Keimbahn antigenunabhängig RAG-1 RAG-2 TdT Lck ZAP-70 CD25 CD44lo HSA später doppelt negativer Thymocyt V-DJ umgeordnet Keimbahn Thymus Prä-T-Rezeptor RAG-1 RAG-2 früher doppelt positiver Thymocyt Prä-T CD4 CD8 HSA V-J umgeordnet T-Zell Rezeptor CD69 CD4 CD8 HSA Lck ZAP-70 später doppelt positiver Thymocyt
Eine Zusammenfassung der Entwicklung menschlicher :β-T-Zellen Marken-protein auf der Oberfläche Umordnungen des -Ketten-Gens Umordnungen des β-Ketten-Gens T-Zellen intra-zelluläre Proteine Naive CD4-T-Zelle CD4 CD62L CD45RA CD5 antigenunabhängig Lck ZAP-70 LKLF CD4-T-Gedächt-niszelle CD4 CD45RO CD44 Lck ZAP-70 Typ 1: IL-5 IL-4 IL-10 Typ 2: IL-2 IFN- CD4 CD45RO CD44hi Fas FasL(Typ1) CD4-T-Effektor-zelle Typ 2 letztlich differenziert zu Peripherie Naive CD8 T-Zelle CD8 CD45RA Antigen-unabhängig CD8-T-Gedächt-niszelle CD8 CD45RO CD44 CD8-T-Effektor-zelle FasL Fas CD8 CD44hi IFN- Granzym Perforin letztlich differenziert zu