Understanding Kinesin-Mediated Active Transport and its Role in Alzheimer's Disease
20 likes | 155 Views
Alzheimer's disease (AD) afflicts over 400,000 individuals in the UK, leading to debilitating cognitive decline. This research investigates the critical role of microtubule active transport in neurons and its disruption in AD. By utilizing computational modeling, we explore kinesin's motion along microtubules to identify modes of failure in active transport. Initial findings reveal complex behaviors such as procession, diffusion, and stasis, shedding light on how alterations in timing affect kinesin’s efficiency. This work aims to deepen our understanding of neurodegeneration processes in AD.

Understanding Kinesin-Mediated Active Transport and its Role in Alzheimer's Disease
E N D
Presentation Transcript
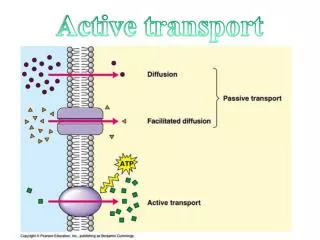
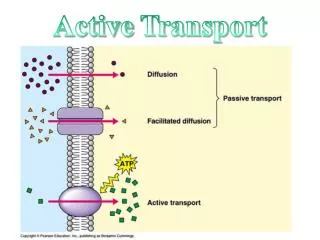
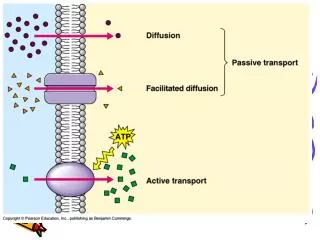
Introduction An estimated 20 million people suffer some form of dementia, over 750,000 in the UK. Of the latter more than 400,000 are afflicted with Alzheimer’s disease (AD), an incurable, fatal condition that progressively destroys the mind over several years. Most cases of AD are late-onset. Why? What is it about ageing neurons that renders them susceptible? One can only speculate but there is a process especially important to neurons as opposed to other types of cell. Failure of microtubule active transport in axons has been proposed as the cause of AD (Gunawardena & Goldstein 2004,2005; Stokin et al 2005). Figure 1 illustrates normal and disrupted axonal active transport. Macromolecules are too large to diffuse through the crowded cytosol and so have to be actively transported to their functional destinations. Axons are long projections emanating from the cell body of the neuron. Microtubule active transport Figure 3 shows kinesin I, a typical motor protein: twin heads (blue) connected via short, flexible necks to a long coiled coil stalk (grey) ending in a fan tail (green). The tail binds the cargo, the heads alternately bind to and detach from the microtubule moving in a head-over-head fashion (figure 2). Figure 1 Disruption of axonal active transport (Gunawardena & Goldstein 2005) In order to investigate active transport, a computational modelling approach has been taken. The initial focus is on the mechanism of motion of conventional kinesin (kinesin I). Figure 2 illustrates the motion of kinesin’s twin motor domains along a microtubule. Results so far indicate 3 possible modes of operation: successful transport, diffusive behaviour, and rigidity. Figure 3 Kinesin (Vale 2003) Modelling microtubule active transport Figure 2 The kinesin walk (Hoenger et al 2000) Richard Wilson • Purpose • The aim of my research is to increase our understanding of active transport and its modes of failure. The focus of this study is the mechanism of normal kinesin procession: how the motor protein walks along the microtubule (MT). • Goals achieved so far: • to show that a leading theoretical model of procession works in silico • to investigate the effects of altering the timing of events in the processionary cycle • to show that the timings that lead to procession in silico match experimental data The computational model Each head is modelled as a finite state machine (FSM) simulated within a 2D box containing a section of microtubule (MT). The maximum possible number of states of the FSM is 8 since a head has 4 states of nucleotide binding (KT, KDP, KD, K0) and is either docked to the microtubule (MT) or not, Not all these states are physically realistic. Experimental evidence indicates that KD has weak affinity for the MT whereas K0, KD and KDP have high affinity for the MT (Asenjo, Krohn & Sosa 2003). Thus the number of states per head can be reduced to 5. Each head follows the same series of states during procession: …KDu -> KD -> K0 -> KT -> KDP -> KDu… The relative timings of these events are parameterised and ranges of timings are simulated to discover the effect of varying the timings on system behaviour. • Results • Initial results reveal a 3 phase system: • diffusion – kinesin fails to process • procession – kinesin walks along the MT • stuck – kinesin stuck in one position on the MT • A linear relationship between timing and phase had been expected but the results appeared chaotic. 2 types of processionary cycle were discovered: symmetric (figure 4) and asymmetric. The latter involves head docking oscillation and is so far resistant to analysis. • Suppressing the asymmetric cycles reduces the system phases to 2: procession or stuck. It also makes the results linear so that a simple timing equality defines procession. Head state key K represents the kinesin head bound to ATP, ADP.P, ADP or 0 (empty nucleotide binding site) and u is appended where a head is undocked Program operation The program is run repeatedly to systematically investigate the effect of altering the timing of state transitions on the motor’s behaviour A simulation run starts with both heads in state KDu not far from the minus end of the MT The motor “diffuses”: constrained random motion is applied to each head until the MT is approached and kinesin engages with the MT The run is terminated when the motor reaches the plus end of the MT or procession fails Figure 4 Symmetric cycle of step time 3. Top row, time; middle row, states of one head; bottom row, states of other head. Docked states with red emphasis, undocked states with green emphasis. • Futures • Analysis of oscillatory behaviour of current model • Introduction of noise into the system to investigate the stability of procession vs degree of stochasticity • Investigation of effects of crowding and temporary blockage • Aim is to increase scope and realism of the model. Prediction The model predicts that procession occurs when the KDP -> KDu transition is delayed relative to the other state transitions. This is in accord with experimental data (Ma & Taylor 1997) who’ve measured a 13 ms undocking time compared to 2 – 7 ms for the timing of the other events for monomeric kinesin. References Asenjo AB, Krohn N & Sosa H (2003) Configuration of the two kinesin motor domains during ATP hydrolysis. Nature Struct Biol10, 836–842 Gunawardena S & Goldstein LS (2004) Cargo-carrying motor vehicles on the neuronal highway: transport pathways and neurodegenerative disease. J Neurobiol58 258-271 Gunawardena S & Goldstein LS (2005) Polyglutamine Diseases and Transport Problems: Deadly Traffic Jams on Neuronal Highways.Arch Neurol62, 46-51 Hoenger A, Thormählen M, Diaz-Avalos R, Doerhoefer M, Goldie KN, Müller J & Mandelkow E (2000) A New Look at the Microtubule Binding Patterns of Dimeric Kinesins. J Mol Biol 297, 1087-1103 Ma YZ & Taylor EW (1997) Kinetic Mechanism of a Monomeric Kinesin Construct. J Biol Chem272 (2), 717–723 Stokin GB, Lillo C, Falzone TL, Brusch RG, Rockenstein E, Mount SL, Raman R, Davies P, Masliah E, Williams DS, et al (2005) Axonopathy and transport deficits early in the pathogenesis of Alzheimer’s disease. Science307, 1282–1288 Vale RD (2003) The Molecular Motor Toolbox for Intracellular Transport. Cell112, 467–480 Yildiz A, Tomishige M, Vale RD & Selvin PR (2004) Kinesin Walks Hand-Over-Hand. Science303 676-8.