Download

1 / 30
300 likes | 358 Views
Dive into the intricate world of membrane transport with details on simple diffusion, carrier-mediated transport, primary and secondary active transport, facilitated diffusion, and ion channels. Learn about transport kinetics, reversible transport, energy requirements, and more.

E N D
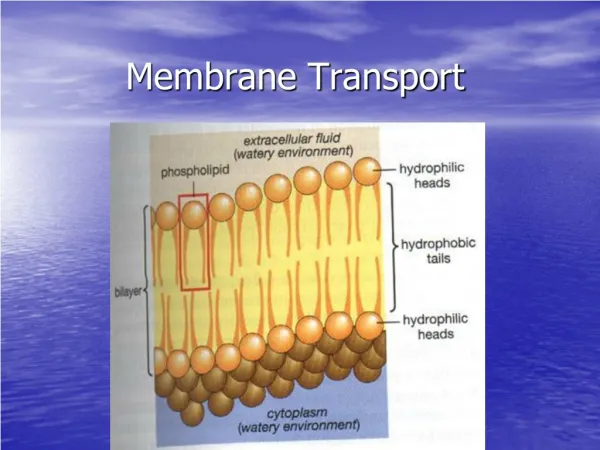
Membrane Transport Jim Huettner
Membrane Flux (moles of solute / sec) • Simple Diffusion • Carrier Mediated Transport • Facilitated Diffusion • Primary Active Transport • Secondary Active Transport • Ion Channels
channel and transporter flux rates co-transport counter-transport Lodish et al., 8th ed. Fig. 11.2
Simple Diffusion • Flux is proportional to external concentration • Flux never saturates Flux [S]o
Carrier-Mediated Transport • Higher flux than predicted by solute permeability • Flux saturates • Binding is selective (D- versus L-forms) • Competition • Kinetics: [S]o << KmÞ M a [S] [S]o = KmÞ M = Mmax / 2 [S]o >> KmÞ M = Mmax Mmax Flux 0.5 Km [S]o
Transport Kinetics k + k - So + Co SCo Si S = Solute C = Carrier dSCo/dt = k + [S]o [C]o – k - [SC]o = 0 at equilibrium Þ k + [S]o [C]o = k - [SC]o k - /k + = ([S]c [C]o)/[SC]o = KmÞ [SC]o = ([S]o [C]o)/Km Fractional Rate = M / Mmax = [SC]o / ([C]o + [SC]o) M = Mmax / (1 + [C]o/[SC]o) = Mmax / (1 + Km/[S]o)
Reversible Transport Co Ci So Si SCo SCi Mnet = Min – Mout = Mmax ) ( 1 1 1 + Km / [S]o 1 + Km / [S]i -
Facilitated Diffusion • Uses bidirectional, symmetric carrier proteins • Flux is always in the directions you expect for simple diffusion • Binding is equivalent on each side of the membrane • Specific Example: Glucose TransportKm = 1.5 mMMmax = 500 µmoles / hr / ml of RBCs [glucose]o = 5 mM [glucose]i = 0.5 mM Mnet = 500 1 1 1 + 1.5 / 5 1 + 1.5 / 0.5 = 500 (0.77 – 0.25) = 260 µmoles glucose / hr (into the cell) ( ) -
Hypothetical Example • Suppose Km(out)¹ Km(in) Km (out) = 1.5 mM Km(in) = 0.1 mM [S]o = 5 mM [S]i = 0.5 mM Mmax = 500 µmoles of S / hr Mnet = 500 1 1 1 + 1.5 / 5 1 + 0.1 / 0.5 = 500 (0.77 – 0.83) = -30 µmoles glucose / hr (out of the cell!) Efflux of S against a concentration gradient would require energy ( ) -
“alternate exposure” Alberts et al., 6th ed.
Primary Active Transport: Driven by ATP I. Class P – all have a phosphorylated intermediate • Na+-K+ ATPase • Ca2+ ATPase • H+-K+ATPase II. ABC transporters III. Class V • H+ transport for intracellular organelles IV. Class F • Synthesize ATP in mitochondria
Primary Active Transport: Na+-K+ ATPase • 3 Na outward / 2 K inward / 1 ATP • Km values: Nain = 20 mM Kout = 2 mM • Blocked by digitalis and ouabain • 2 subunits, beta and alpha (the pump) • Turnover = 300 Na+ / sec / pump site @ 37 °C 3 Na out ATP ADP + Pi in 2 K
Na+-K+ ATPase: Operation of the pump • 3 Na+ bind on the intracellular face of the alpha subunit • Mg-ATP binds and Pi is transferred to an aspartate on the alpha subunit • The pump changes conformation and releases Na+ to the extracellular face • 2 K+ bind from outside • The aspartate-phosphate bond is broken and the pump returns to release K+ on the inside • If the energy for each pump cycle is supplied by ATP, can we be sure that ATP has enough energy to do it?
Secondary Active Transport • Energy stored in the Na+ gradient is used to power the transport of a variety of solutes Co-transport:glucose, amino acids and other molecules are pumped in Counter-transport:Ca2+ or H+ are pumped out 2 or 3 Na+ / 1 Ca2+ ; 1 Na+ / 1 H+ • These transport proteins do not hydrolyze ATP directly; but they work at the expense of the Na+ gradient which must be maintained by the Na-K ATPase
Energy available from ATP H2O ATP ADP + Pi DG = Gproducts – G reactants Chemical Energy (G) = RT ln [C] DG = DG° + 2.3 RT (log ([ADP] [Pi]) – log [ATP]) 2.3 RT = 5.6 kiloJoules / mole @ 20° C DG° = -30 kiloJoules / mole @ 20° C, pH 7.0 and 1 Molar [reactants] and [products] “Standard Conditions”
Energy Depends on Concentrations DG = -30 – 5.6 log [ATP] kJ / mole [ADP] [Pi] • The energy available per molecule of ATP depends on: • [ATP] @ 4mM, [ADP] @ 400 µM, [Pi] @ 2 mM • Þ per mole of ATP hydrolyzed: • DG = -30 kJ – 5.6 kJ * log 4 x 10-3 • 2 x 10-3 * 4 x 10-4 • = -30 kJ - 21 kJ = -51 kiloJoules per mole of ATP • How much energy from a single molecule of ATP?
Energy per molecule of ATP We need a new unit of energy – the milli-electron-Volt Volt = Joule / Coulomb Þ 1 Joule = 1 Volt * 1 Coulomb Define an “Electron –Volt” (eV) as the energy of 1 electron at a potential of 1 Volt and a milli-electron-Volt (meV) as the energy of 1 electron at a potential of 1 milliVolt Charge of a single electron = 1.602 x 10-19 Coulombs 1 Joule = 1 V * (1000 mV / V) * 1 Coulomb = 6.242 x 1021 meV 1.602 x 10-19 Coulombs / electron Energy per molecule: -51 x 103 J / mole * 6.242 x1021 meV / J = -530 meV / molecule 6.02 x 1023 molecules / mole
Review of this Calculation • DG = Gproducts – Greactants Chemical Energy (G) = RT ln [C] • DG = -30 – 5.6 * log [ATP] kJ / mole • [ADP] [Pi] • = -51 kJ / mole = -530 meV / molecule • The negative sign means energy is released when ATP is broken downs • The energy available per molecule of ATP depends on: [ATP], [ADP], and [Pi] • Conclusion: Cells can obtain at least 500 meV per molecule of ATP
Energy in the Sodium Gradient Consider Na+ movement from outside to inside: DG = Gproducts – Greactants = Ginside – Goutside DGtotal = DGelectrical + DGchemical Conditions for our sample calculation: Vm = -60 mV [Na+]out = 140 mM [Na+]in = 14 mM and 2.3 RT = 60 meV / molecule
Energy in the Na Gradient: Electrical Term • DGelectrical = e * mVin – e * mVout • = +1e * -60 mV – (+1e) * 0 mV • = -60 meV • negative sign means energy is released moving from outside to inside • 60 meV is the energy required to move a charged ion (z=1) up a voltage gradient of 60 mV (assuming zero concentration gradient)
Energy in the Na Gradient: Chemical Term • DGchemical = 2.3 RT (log [Na+]in – log [Na+]out) • = 60 meV * (-1) • = -60 meV • negative sign means energy is released moving from outside to inside • 60 meV is the energy required to move a molecule up a 10 fold concentration gradient (true for an uncharged molecule or for a charged molecule when there is no voltage gradient)
Energy in the Na Gradient: Total • DGtotal = DGelectrical + DGchemical = -120 meV • 120 milli-electron-Volts of energy would be required to pump a single Na+ ion out of the cell up a 10 fold concentration gradient and a 60 mV voltage gradient. • Hydrolysis of a single ATP molecule can provide at least 500 meV of energy – enough to pump 4 Na+ ions. • A single Na+ ion moving from outside to inside would be able to provide 120 meV of energy, which could be used to pump some other molecule, such as glucose, an amino acid, Ca2+ or H+ up a concentration gradient
First Example: Na+ / Ca2+ exchange Compare the internal [Ca2+] for exchange ratios of 2 Na+ : 1 Ca2+v.s. 3 Na+ : 1 Ca2+ Vm = -60 mV, [Ca2+]out = 1.5 mM [Ca2+]in = ? Ca2+ moves from inside to outside DG = Gproducts – Greactants = Goutside – Ginside DGelectrical = (+2e) * (0 mV) – (+2e) * (-60 mV) = +120 meV DGchemical = 60 meV (log 1.5 – log ?)
Na+ / Ca2+ exchange DGtotal = DGE + DGC = 120 meV + 60 meV log (1.5 / ?) 2 Na+Þ 240 meV 240 = 120 + 60 log (1.5 / ?) 120 / 60 = log (1.5 / ?) 102 = 1.5 / ? ? = 15 µM 3 Na+Þ 360 meV 360 = 120 + 60 log (1.5 / ?) 240 / 60 = log (1.5 / ?) 104 = 1.5 / ? ? = 0.15 µM Internal [Ca2+]can be reduced 100 fold lowerfor 3 Na : 1 Cav.s. 2 Na : 1 Ca
Second Example: Acid Secretion Blood Cytoplasm Stomach Lumen 0 mV -70 mV ? ** -70 mV [K] = 145 mM [K] = 10 mM pH = 7 pH = 1 H ATP ADP + Pi ** Schreiber et al. Am J Physiol Gast Liver Phys279:597-604 (2000) K H+ moves from inside to outside DG = Gproducts – Greactants = Goutside – Ginside DGelectrical = (+1e) * (-70 mV) – (+1e) * (-70 mV) = 0 meV DGchemical = 60 meV (-1 – (-7)) = +360 meV per proton remember pH = -log [H+]
Acid Secretion (continued) K+ moves from outside to inside DG = Gproducts – Greactants = Ginside - Goutside DGelectrical = (+1e) * (-70 mV) – (+1e) * (-70 mV) = 0 meV DGchemical = 60 meV (log 145 – log 10) = +69.7 meV per K DGtotal = DGproton + DGpotassium = +429.7 meV Since 1 molecule of ATP can supply ~ 500 meV, it is unlikely that more than one proton is pumped per cycle
Summary: Energetics • Transport Energetics • A molecule of ATP donates about 500 meV • It takes 60 meV to transport up a 60 mV electrical gradient • It takes 60 meV to transport up a 10 fold concentration gradient • A single sodium ion donates approximately 120 meV • Acid Secretion • Parietal cells in the stomach secrete HCl • The H-K ATPase in the lumenal membrane exchanges 1 H+ for 1 K+ per ATP
Biological Control of Secretion • Cephalic Phase – From CNS via vagus nerve • Gastric Phase – A reflex response to stomach distention • Secretory agents: • Acetylcholine – released from neurons intrinsic to the gut. Elevates intracellular calcium. Muscarinic receptors. • Gastrin – small peptide released into the circulation from cells near the pylorus. Elevated intracellular calcium. • Histamine – continuously present in background concentrations.No evidence for elevation during secretion. H2 receptors on parietal cells stimulate Adenylate cyclase. Synergistic with ACh and Gastrin. • It is not yet known how these agents lead to activation of parietal cells