Download
1 / 77
780 likes | 934 Views
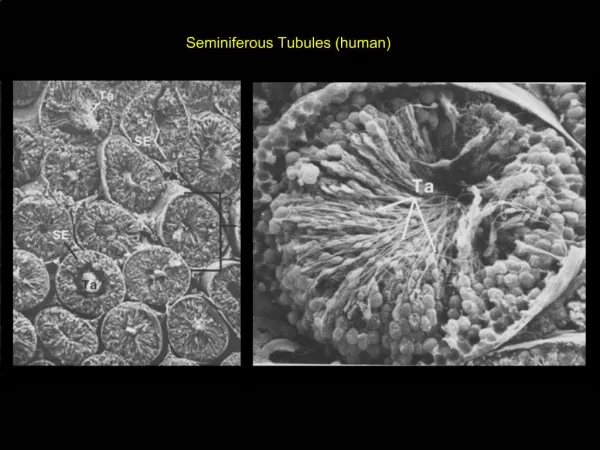
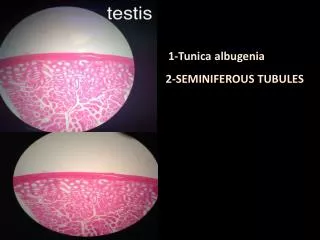
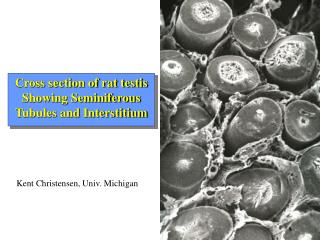
Cross section of rat testis Showing Seminiferous Tubules and Interstitium. Kent Christensen, Univ. Michigan. Functional and Anatomical Compartments of the Testis.

E N D
Cross section of rat testisShowing Seminiferous Tubules and Interstitium Kent Christensen, Univ. Michigan
Interstitium of rat testis showing endothelium, Leydig cells (L), and macrophages (arrow). Note close association of macrophages and Leydig cells. Scott Miller, Univ Utah
Close association of Leydig cell and macrophage, lower panel shows close up of “digitation” of Leydig cell process extending onto macrophage surface. Scott Miller, Univ. Utah
Macrophage-Leydig cell interactions Cytokines, ROS ?
LH Extracellularlipoprotein Cholesterolpool acetate ATP cAMP cholesterol PKA+ DYm Pregnenolone ++ Transcription 3bHSD Progesterone P450c17 Androstenedione 17bHSD TESTOSTERONE
P450c17 is sensitive to transcriptional repression • Of all the steroidogenic enzymes, P450c17 is the most sensitive to repression • Most cytokines tested inhibit c17 transcription: • IL-1a/b, IL-2, IL-6, TNFa, TGFb, INFa/b, INFg • Inflammatory mediators: PGF2a, ceramide, vasopressin, PKC agonists • Environmental disruptors such as dioxin, pthalates, PAHs, etc. are inhibitory • Androgen-mediated feedback repression
IL-1, TNFa and PMA vs. steroidogenic mRNA expression P450scc P450c17 +IL-1 cAMP +PMA +TNFa
TNFa and PMA stimulate translocation of PKCa from cytoplasm to membrane control No antibody PMA TNFa
Comparison of cAMP-responsive sequences in mouse and rat Binding sites (ATF2/cjun-like,Steroidogenic factor 1,StF-IT-2, andStF-IT-1/COUP-TF1) are shown in bold color in the sequence for the species in which it was identified. Those that are conserved between species are underlined. Sequence differences are shown in blue for mouse and pink for rat.
Characterization of the Cyp17 Promoter Revealed a Region Between -245 and -346 Responsible for the Minimal cAMP Responsiveness of the Gene
Site-directed mutagenesis of Cyp17 CRR (-346 to –245) • Oligos were designed to place an XhoI once every ten base pairs within the 100 base pair CRR. • This resulted in changing as few as three (mutant 6) to as many as six (mutant 1 and 7) of every ten nucleotides. • Mutagenesis was performed with Altered Sites (Promega) and all mutants were verified by sequencing.
Putative sites revealed by mutants gcaacctgatgacattaatt attaactgtg cagcacttttgacattacag CTCGAGtgat CTcGAGaatt CtCGaGtgtg cTCGaGtttt CTcGAGacag mut 1 mut 2 mut 3 mut 4 mut 5 cacgcactctgaaaccttga tcttaatctg atagcatttg cctctgggag cTcgAGctct CTCGAGttga CTCGaGtctg CtCgAGtttg cACGAgggag mut 6 mut 7 mut 8 mut 9 mut 10 ATF2 AhR/Arnt(core sequence) SF-1
ATF2 mutants 2,5,9 C/EBPb– upstream site AhR/ARNT mutant 6 SF-1 mutant 7 ARE Putative regulatory motifs revealed by mutagenesis ? -250 -440
The Minimal cAMP Responsive Region of the Cyp17 Promoter (CRR): -346 TGATGACATTAATTATTAA CTGTGCAGCA CTTTTGACATTACAGCACGC ACTCTGAAAC CTTGATCTTA ATCTAGCATTTGCCTCTGGG AGGATCCATA GCG -245 -346 -245 Putative ATF-2 binding site
Binding of Nuclear Proteins to the CRR Probe is Augmented by Treatment of MA-10 Cells with cAMP
Nuclear Proteins from Primary Leydig Cells Form Two Complexes with the CRR Probe
The Upstream ATF-2 C/EBPb Binding Site -450 TTGTGTGACCTTATGCAAAC TAACCCA -423 -245 -450
Nuclear Proteins from Control and cAMP-Treated MA-10 cells Bind to the Upstream ATF-2 C/EBPb Probe
Incubation of the Upstream ATF-2 C/EBPb probe with Nuclear Proteins Isolated from Primary Leydig Cells Results in Complex Formation
Formation of the Higher Order Complex Formed by the CRR Complex is Decreased by Addition of Unlabeled ATF-2 or C/EBPb Competitor Oligos
Binding of Nuclear Proteins to the Upstream ATF-2 C/EBPb Probe can be Inhibited by Addition of ATF-2 or C/EBPb Competitor Oligos
Overexpression of C/EBPb Induces Transcription of the -491/-255 Cyp 17 reporter
ATF-2 Expression in MA-10 Cells is Not Affected by cAMP Treatment
C/EBPb Expression is Significantly Increased in MA-10 Cells with cAMP p38 p34 **
Summary of Cyp17 study • TNFa-mediated inhibition of transcription involves activation of PKC • ATF2 and C/EBPb participate cooperatively in cAMP-induction of transcription • ATF2 is constitutively expressed • C/EBPb expression is induced by cAMP
Hypotheses • ATF2 and C/EBPb interact as heterodimers binding to the “ATF2” sites in the promoter • The stoichiometry of C/EBPb and ATF2 interaction is critical to driving transcription • Repressors may act by inhibiting C/EBPb expression or through post-translational modifications that inhibit its activity • C/EBPb phosphorylation by PKC may block it from interacting with ATF2
P450scc P450c17 3b-HSD actin - + - + - + - + - + LPS 2h 4h 6h 8h 24h time Effect of LPS on steroidogenic mRNA levels
LPS vs. serum testosterone: 2-24 hours control 14 LPS 12 10 8 Testosterone (ng/ml) 6 4 2 0 2 h 4 h 6 h 8 h 24 h Time post LPS
Steroidogenic Acute Regulatory Protein: StAR • Essential for steroid hormone biosynthesis • Cyclic-AMP dependent expression • Facilitates cholesterol transfer across inner-mitochondrial (aqueous) space • Translated as a 37 kDa precursor protein that is processed to the 30 kDa mature form as it translocates into the mitochondria • Cholesterol transport activity depends on intact DYm
N' 32 kDa N' 30 kDa 37 StAR Processing 32 30 Inner-mitochondrial forms Cytosol 37 kDa N' cholesterol transfer critical region signal peptides Outer mitochondrial membrane Inner- mitochondrial membrane matrix
StAR N-terminal localization expression clones MTS 1-37 ITS 38-47 pCMV-StAR TAA StAR-stop MTS 1-37 StAR D-ITS StAR D-N47 Tom20 OMTS StAR/Tom20 CCHL IMSS StAR/CCHL
What mediates the acute LPS inhibition? • Tested numerous inflammatory mediators in Leydig cells in vitro-- none mimicked the acute LPS “effect” • cytokines (TNFa, IL-1, IL-6, IFNg, TGFb) • prostaglandins (PGF2a, PGE) • catecholamines (norepi, isoproteranol) • ceramide (C2, C8) • Most nitric oxide donors (Sin-1, SNAP, SNP, Nor-3) • Calcium inophore (A23187)
LPS vs. StAR protein expression: 2 hr after injection 37 kDa 30 kDa con LPS
Carbonyl cyanide m-chlorophenylhydrazone (cccp) • Carbonyl cyanide m-chlorophenyl-hydrazone (cccp): potent uncoupler of oxidative phosphorylation; protonophore, mitochondrial disrupter. • Causes transient disruption of DYm
H+ DYm e- Mitochondrial respiration, OX-PHOS and DYm
Effect of CCCP on StAR protein 37 kDa 30 kDa Control cAMP cAMP + cccp cccp