Download
1 / 57
570 likes | 941 Views
DNA Sequencing. DNA Sequencing – gel electrophoresis. Start at primer (restriction site) Grow DNA chain Include dideoxynucleoside (modified a, c, g, t) Stops reaction at all possible points Separate products with length, using gel electrophoresis. Electrophoresis diagrams.

E N D
DNA Sequencing – gel electrophoresis • Start at primer (restriction site) • Grow DNA chain • Include dideoxynucleoside (modified a, c, g, t) • Stops reaction at all possible points • Separate products with length, using gel electrophoresis
Method to sequence longer regions genomic segment cut many times at random (Shotgun) Get one or two reads from each segment ~500 bp ~500 bp
Reconstructing the Sequence (Fragment Assembly) reads Cover region with ~7-fold redundancy (7X) Overlap reads and extend to reconstruct the original genomic region
Definition of Coverage C Length of genomic segment: L Number of reads: n Length of each read: l Definition:Coverage C = n l / L How much coverage is enough? Lander-Waterman model: Assuming uniform distribution of reads, C=10 results in 1 gapped region /1,000,000 nucleotides
Repeats Bacterial genomes: 5% Mammals: 50% Repeat types: • Low-Complexity DNA (e.g. ATATATATACATA…) • Microsatellite repeats (a1…ak)N where k ~ 3-6 (e.g. CAGCAGTAGCAGCACCAG) • Transposons • SINE(Short Interspersed Nuclear Elements) e.g., ALU: ~300-long, 106 copies • LINE(Long Interspersed Nuclear Elements) ~4000-long, 200,000 copies • LTRretroposons(Long Terminal Repeats (~700 bp) at each end) cousins of HIV • Gene Families genes duplicate & then diverge (paralogs) • Recent duplications ~100,000-long, very similar copies
AGTAGCACAGACTACGACGAGACGATCGTGCGAGCGACGGCGTAGTGTGCTGTACTGTCGTGTGTGTGTACTCTCCTAGTAGCACAGACTACGACGAGACGATCGTGCGAGCGACGGCGTAGTGTGCTGTACTGTCGTGTGTGTGTACTCTCCT Sequencing and Fragment Assembly 3x109 nucleotides 50% of human DNA is composed of repeats Error! Glued together two distant regions
What can we do about repeats? Two main approaches: • Cluster the reads • Link the reads
What can we do about repeats? Two main approaches: • Cluster the reads • Link the reads
What can we do about repeats? Two main approaches: • Cluster the reads • Link the reads
AGTAGCACAGACTACGACGAGACGATCGTGCGAGCGACGGCGTAGTGTGCTGTACTGTCGTGTGTGTGTACTCTCCTAGTAGCACAGACTACGACGAGACGATCGTGCGAGCGACGGCGTAGTGTGCTGTACTGTCGTGTGTGTGTACTCTCCT A R B D R C Sequencing and Fragment Assembly 3x109 nucleotides ARB, CRD or ARD, CRB ?
AGTAGCACAGACTACGACGAGACGATCGTGCGAGCGACGGCGTAGTGTGCTGTACTGTCGTGTGTGTGTACTCTCCTAGTAGCACAGACTACGACGAGACGATCGTGCGAGCGACGGCGTAGTGTGCTGTACTGTCGTGTGTGTGTACTCTCCT Sequencing and Fragment Assembly 3x109 nucleotides
Strategies for whole-genome sequencing • Hierarchical – Clone-by-clone • Break genome into many long pieces • Map each long piece onto the genome • Sequence each piece with shotgun Example: Yeast, Worm, Human, Rat • Online version of (1) – Walking • Break genome into many long pieces • Start sequencing each piece with shotgun • Construct map as you go Example: Rice genome • Whole genome shotgun One large shotgun pass on the whole genome Example: Drosophila, Human (Celera), Neurospora, Mouse, Rat, Dog
a BAC clone map Hierarchical Sequencing Strategy • Obtain a large collection of BAC clones • Map them onto the genome (Physical Mapping) • Select a minimum tiling path • Sequence each clone in the path with shotgun • Assemble • Put everything together genome
Methods of physical mapping Goal: Make a map of the locations of each clone relative to one another Use the map to select a minimal set of clones to sequence Methods: • Hybridization • Digestion
1. Hybridization Short words, the probes, attach to complementary words • Construct many probes • Treat each BAC with all probes • Record which ones attach to it • Same words attaching to BACS X, Y overlap p1 pn
Hybridization – Computational Challenge p1p2 …………………….pm 0 0 1 …………………..1 Matrix: m probes n clones (i, j): 1, if pi hybridizes to Cj 0, otherwise Definition: Consecutive ones matrix 1s are consecutive in each row & col Computational problem: Reorder the probes so that matrix is in consecutive-ones form Can be solved in O(m3) time (m > n) C1C2 ……………….Cn 1 1 0 …………………..0 1 0 1…………………...0 pi1pi2…………………….pim 1 1 1 0 0 0……………..0 0 1 1 1 1 1……………..0 0 0 1 1 1 0……………..0 Cj1Cj2 ……………….Cjn 0 0 0 0 0 0………1 1 1 0 0 0 0 0 0 0………0 1 1 1
Hybridization – Computational Challenge pi1pi2………………………………….pim pi1pi2…………………….pim If we put the matrix in consecutive-ones form, then we can deduce the order of the clones & which pairs of clones overlap 1 1 1 0 0 0……………..0 0 1 1 1 1 1……………..0 0 0 1 1 1 0……………..0 Cj1Cj2 ……………….Cjn Cj1Cj2 ……………….Cjn 0 0 0 0 0 0………1 1 1 0 0 0 0 0 0 0………0 1 1 1
Hybridization – Computational Challenge p1p2 …………………….pm 0 0 1 …………………..1 Additional challenge: A probe (short word) can hybridize in many places in the genome Computational Problem: Find the order of probes that implies the minimal probe repetition Equivalent: find the shortest string of probes such that each clone appears as a substring APX-hard Solutions: Greedy, probabilistic, lots of manual curation C1C2 ……………….Cn 1 1 0 …………………..0 1 0 1…………………...0
2. Digestion Restriction enzymes cut DNA where specific words appear • Cut each clone separately with an enzyme • Run fragments on a gel and measure length • Clones Ca, Cb have fragments of length { li, lj, lk } overlap Double digestion: Cut with enzyme A, enzyme B, then enzymes A + B
The Walking Method • Build a very redundant library of BACs with sequenced clone-ends (cheap to build) • Sequence some “seed” clones • “Walk” from seeds using clone-ends to pick library clones that extend left & right
Walking off a Single Seed • Low redundant sequencing • Many sequential steps
Walking off a single clone is impractical • Cycle time to process one clone: 1-2 months • Grow clone • Prepare & Shear DNA • Prepare shotgun library & perform shotgun • Assemble in a computer • Close remaining gaps • A mammalian genome would need 15,000 walking steps !
Walking off several seeds in parallel • Few sequential steps • Additional redundant sequencing In general, can sequence a genome in ~5 walking steps, with <20% redundant sequencing Efficient Inefficient
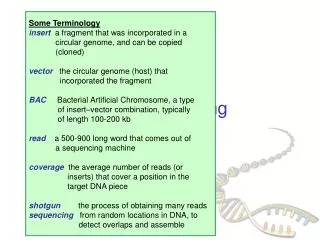
Some Terminology insert a fragment that was incorporated in a circular genome, and can be copied (cloned) vector the circular genome (host) that incorporated the fragment BACBacterial Artificial Chromosome, a type of insert–vector combination, typically of length 100-200 kb read a 500-900 long word that comes out of a sequencing machine coveragethe average number of reads (or inserts) that cover a position in the target DNA piece shotgun the process of obtaining many reads sequencing from random locations in DNA, to detect overlaps and assemble
cut many times at random Whole Genome Shotgun Sequencing genome plasmids (2 – 10 Kbp) forward-reverse paired reads known dist cosmids (40 Kbp) ~500 bp ~500 bp
Fragment Assembly Given N reads… Where N ~ 30 million… We need to use a linear-time algorithm
Steps to Assemble a Genome Some Terminology read a 500-900 long word that comes out of sequencer mate pair a pair of reads from two ends of the same insert fragment contig a contiguous sequence formed by several overlapping reads with no gaps supercontig an ordered and oriented set (scaffold) of contigs, usually by mate pairs consensus sequence derived from the sequene multiple alignment of reads in a contig 1. Find overlapping reads 2. Merge some “good” pairs of reads into longer contigs 3. Link contigs to form supercontigs 4. Derive consensus sequence ..ACGATTACAATAGGTT..
1. Find Overlapping Reads (read, pos., word, orient.) aaactgcag aactgcagt actgcagta … gtacggatc tacggatct gggcccaaa ggcccaaac gcccaaact … actgcagta ctgcagtac gtacggatc tacggatct acggatcta … ctactacac tactacaca (word, read, orient., pos.) aaactgcag aactgcagt acggatcta actgcagta actgcagta cccaaactg cggatctac ctactacac ctgcagtac ctgcagtac gcccaaact ggcccaaac gggcccaaa gtacggatc gtacggatc tacggatct tacggatct tactacaca aaactgcagtacggatct aaactgcag aactgcagt … gtacggatct tacggatct gggcccaaactgcagtac gggcccaaa ggcccaaac … actgcagta ctgcagtac gtacggatctactacaca gtacggatc tacggatct … ctactacac tactacaca
T GA TACA | || || TAGA TAGT 1. Find Overlapping Reads • Find pairs of reads sharing a k-mer, k ~ 24 • Extend to full alignment – throw away if not >98% similar TAGATTACACAGATTAC ||||||||||||||||| TAGATTACACAGATTAC • Caveat: repeats • A k-mer that occurs N times, causes O(N2) read/read comparisons • ALU k-mers could cause up to 1,000,0002 comparisons • Solution: • Discard all k-mers that occur “too often” • Set cutoff to balance sensitivity/speed tradeoff, according to genome at hand and computing resources available
1. Find Overlapping Reads Create local multiple alignments from the overlapping reads TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAG TTACACAGATTATTGA TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAG TTACACAGATTATTGA TAGATTACACAGATTACTGA
1. Find Overlapping Reads • Correcterrors using multiple alignment TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAGATTACACAGATTATTGA TAG-TTACACAGATTATTGA TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAG-TTACACAGATTACTGA TAG-TTACACAGATTATTGA insert A correlated errors— probably caused by repeats disentangle overlaps replace T with C TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA TAGATTACACAGATTACTGA In practice, error correction removes up to 98% of the errors TAG-TTACACAGATTATTGA TAG-TTACACAGATTATTGA
2. Merge Reads into Contigs • Overlap graph: • Nodes: reads r1…..rn • Edges: overlaps (ri, rj, shift, orientation, score) Reads that come from two regions of the genome (blue and red) that contain the same repeat Note: of course, we don’t know the “color” of these nodes
repeat region 2. Merge Reads into Contigs We want to merge reads up to potential repeat boundaries Unique Contig Overcollapsed Contig
repeat region 2. Merge Reads into Contigs • Ignore non-maximal reads • Merge only maximal reads into contigs
2. Merge Reads into Contigs • Remove transitively inferable overlaps • If read r overlaps to the right reads r1, r2, and r1 overlaps r2, then (r, r2) can be inferred by (r, r1) and (r1, r2) r r1 r2 r3
2. Merge Reads into Contigs repeat boundary??? sequencing error • Ignore “hanging” reads, when detecting repeat boundaries b a … b a
Overlap graph after forming contigs Unitigs: Gene Myers, 95
Repeats, errors, and contig lengths • Repeats shorter than read length are easily resolved • Read that spans across a repeat disambiguates order of flanking regions • Repeats with more base pair diffs than sequencing error rate are OK • We throw overlaps between two reads in different copies of the repeat • To make the genome appear less repetitive, try to: • Increase read length • Decrease sequencing error rate Role of error correction: Discards up to 98% of single-letter sequencing errors decreases error rate decreases effective repeat content increases contig length
2. Merge Reads into Contigs • Insert non-maximal reads whenever unambiguous
3. Link Contigs into Supercontigs Normal density Too dense Overcollapsed Inconsistent links Overcollapsed?
3. Link Contigs into Supercontigs Find all links between unique contigs Connect contigs incrementally, if 2 links supercontig (aka scaffold)
3. Link Contigs into Supercontigs Fill gaps in supercontigs with paths of repeat contigs
4. Derive Consensus Sequence TAGATTACACAGATTACTGA TTGATGGCGTAA CTA Derive multiple alignment from pairwise read alignments TAGATTACACAGATTACTGACTTGATGGCGTAAACTA TAG TTACACAGATTATTGACTTCATGGCGTAA CTA TAGATTACACAGATTACTGACTTGATGGCGTAA CTA TAGATTACACAGATTACTGACTTGATGGGGTAA CTA TAGATTACACAGATTACTGACTTGATGGCGTAA CTA Derive each consensus base by weighted voting (Alternative: take maximum-quality letter)