Download
1 / 1
10 likes | 65 Views
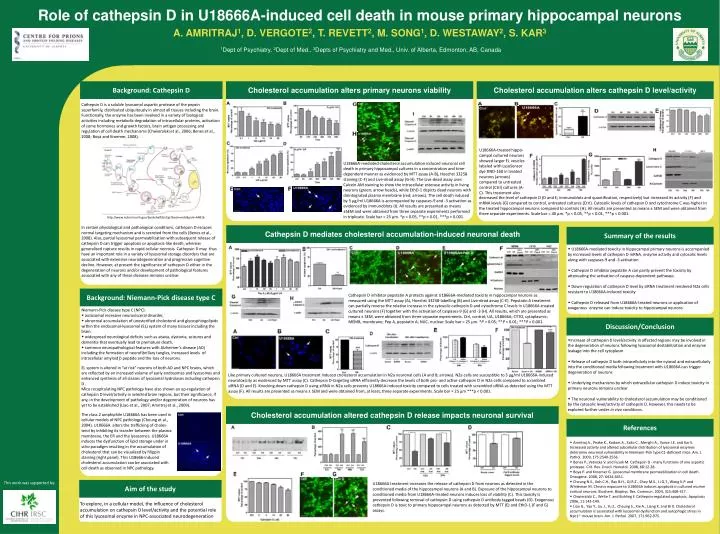
Role of cathepsin D in U18666A-induced cell death in mouse primary hippocampal neurons A. AMRITRAJ 1 , D. VERGOTE 2 , T. REVETT 2 , M. SONG 1 , D. WESTAWAY 2 , S. KAR 3 1 Dept of Psychiatry, 2 Dept of Med., 3 Depts of Psychiatry and Med., Univ. of Alberta, Edmonton, AB, Canada.

E N D
Role of cathepsin D in U18666A-induced cell death in mouse primary hippocampal neurons A. AMRITRAJ1, D. VERGOTE2, T. REVETT2, M. SONG1, D. WESTAWAY2, S. KAR31Dept of Psychiatry, 2Dept of Med., 3Depts of Psychiatry and Med., Univ. of Alberta, Edmonton, AB, Canada Background: Cathepsin D Cholesterol accumulation alters primary neurons viability Cholesterol accumulation alters cathepsin D level/activity Cathepsin D is a soluble lysosomal aspartic protease of the pepsin superfamily, distributed ubiquitously in almost all tissues including the brain. Functionally, the enzyme has been involved in a variety of biological activities including metabolic degradation of intracellular proteins, activation of some hormones and growth factors, brain antigen processing and regulation of cell death mechanisms (Chwieralskiet al., 2006; Benes et al., 2008; Boya and Kroemer, 2008). U18666A-treated hippo-campal cultured neurons showed larger EL vesicles labeled with LysoSensor dye DND-160 in treated neurons (arrows) compared to untreated control (Ctrl) cultures (A-C). This treatment also U18666A-mediated cholesterol accumulation induced neuronal cell death in primary hippocampal cultures in a concentration and time-dependent manner as evidenced by MTT assay (A-B), Hoechst 33258 staining (C-F) and Live-dead assay (G-H). The Live-dead assay uses Calcein AM staining to show the intracellular esterase activity in living neurons (green; arrow heads), while EthD-1 depicts dead neurons with disintegrated plasma membrane (red; arrows). The cell death induced by 5 μg/ml U18666A is accompanied by caspases-9 and -3 activation as evidenced by immunoblots (I). All results are presented as means ±SEM and were obtained from three separate experiments performed in triplicate. Scale bar = 25 µm. *p < 0.05, **p < 0.01, ***p < 0.001. decreased the level ofcathepsin D (D and E; immunoblots and quantification, respectively) but increased its activity (F) and mRNA levels (G) compared to control, untreated cultures (Ctrl). Cytosolic levels of cathepsin D and cytochrome C was higher in the treated hippocampal neurons compared to controls (H). All results are presented as means ± SEM and were obtained from three separate experiments. Scale bar = 40 µm. *p < 0.05, **p < 0.01, ***p < 0.001. http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=mcb&part=A4816 In certain physiological and pathological conditions, cathepsin D escapes normal targeting mechanism and is secreted from the cells (Benes et al., 2008). Also, partial lysosomalpermeabilizationwith subsequent release of cathepsin D can trigger apoptosis or apoptosis-like death, whereas generalized rupture results in rapid cellular necrosis. Cathepsin D may thus have an important role in a variety of lysosomal storage disorders that are associated with extensive neurodegenerationand progressive cognitive decline. However, at present the significance of cathepsin D either in the degeneration of neurons and/or development of pathological features associated with any of these diseases remains unclear. Cathepsin D mediates cholesterol accumulation-induced neuronal death Summary of the results • U18666A-mediated toxicity in hippocampal primary neurons is accompanied by increased levels of cathepsin D mRNA, enzyme activity and cytosolic levels along with caspases-9 and -3 activation • Cathepsin D inhibitor pepstatin A can partly prevent the toxicity by attenuating the activation of caspase-dependent pathways • Down-regulation of cathepsin D level by siRNA treatment rendered N2a cells resistant to U18666A-induced toxicity • Cathepsin D released from U18666A-treated neurons or application of exogenous enzyme can induce toxicity to hippocampal neurons Cathepsin D inhibitor pepstatin A protects against U18666A-mediated toxicity in hippocampal neurons as measured using the MTT assay (A), Hoechst 33258-labelling (B) and Live-dead assay (C-E). Pepstatin A treatment can partially reverse the relative increase in the cytosoliccathepsin D and cytochrome C levels in U18666A-treated cultured neurons (F) together with the activation of caspases-9 (G) and -3 (H). All results, which are presented as means ± SEM, were obtained from three separate experiments. Ctrl, control; UA, U18666A; CYTO, cytoplasmic; MEMB, membrane; Pep A, pepstatin A; NUC, nuclear. Scale bar = 25 µm. *P < 0.05; **P < 0.01; ***P < 0.001. Background: Niemann-Pick disease type C • Niemann-Pick disease type C (NPC): • autosomalrecessive neurovisceraldisorder, • abnormal accumulation of unesterified cholesterol and glycosphingolipids within the endosomal-lysosomal (EL) system of many tissues including the brain. • widespread neurological deficits such as ataxia, dystonia, seizures and dementia that eventually lead to premature death, • common neuropathological features with Alzheimer’s disease (AD) including the formation of neurofibrillary tangles, increased levels of intracellular amyloidbpeptide and the loss of neurons. Discussion/Conclusion • Increase of cathepsin D level/activity in affected regions may be involved in the degeneration of neurons following lysosomal destabilization and enzyme leakage into the cell cytoplasm • Release of cathepsin D both intracellularly into the cytosol and extracellularly into the conditioned media following treatment with U18666A can trigger degeneration of neurons • Underlying mechanisms by which extracellular cathepsin D induce toxicity in primary neurons remains unclear • The neuronal vulnerability to cholesterol accumulation may be conditioned by the cytosolic level/activity of cathepsin D. However, this needs to be explored further under in vivo conditions. EL system is altered in “at risk” neurons of both AD and NPC brains, which are reflected by an increased volume of early endosomes and lysosomes and enhanced synthesis of all classes of lysosomalhydrolases including cathepsinD . Mice recapitulating NPC pathology have also shown an up-regulation of cathepsin D level/activity in selected brain regions, but their significance, if any, in the development of pathology and/or degeneration of neurons has yet to be established (Liao et al., 2007; Amritraj et al., 2009). Like primary cultured neurons, U18666A treatment induced cholesterol accumulation in N2a neuronal cells (A and B; arrows). N2a cells are susceptible to 3 μg/ml U18666A-induced neurotoxicity as evidenced by MTT assay (C). Cathepsin D-targeting siRNA efficiently decrease the levels of both pro- and active-cathepsin D in N2a cells compared to scrambled siRNA (D and E). Knocking down cathepsin D using siRNA in N2a cells prevents U18666A-induced toxicity compared to cells treated with scrambled siRNA as detected using the MTT assay (F). All results are presented as means ± SEM and were obtained from, at least, three separate experiments. Scale bar = 25 µm ***p < 0.001. Aim of the study To explore, in a cellular model, the influence of cholesterol accumulation on cathepsin D level/activity and the potential role of this lysosomal enzyme in NPC-associated neurodegeneration Cholesterol accumulation altered cathepsin D release impacts neuronal survival References The class 2 amphiphile U18666A has been used in cellular models of NPC pathology (Cheung et al., 2004). U18666A alters the trafficking of choles-terol by inhibiting its transfer between the plasma membrane, the ER and the lysosomes. U18666A induces the dysfunction of lipid storage under in vitro paradigm resulting in the accumulation of cholesterol that can be visualized by filippin staining (right panel). This U18666-induced cholesterol accumulation can be associated with cell death as observed in NPC pathology. Amritraj A., Peake K., Kodam A., Salio C., Merighi A., Vance J.E. and Kar S. Increased activity and altered subcellular distribution of lysosomal enzymes determine neuronal vulnerability in Niemann-Pick type C1-deficient mice. Am. J. Pathol. 2009, 175:2540-2556. Benes P., Vetvicka V. and Fusek M. Cathepsin D - many functions of one aspartic protease. Crit. Rev. Oncol. Hematol. 2008, 68:12-28. Boya P. and Kroemer G. Lysosomal membrane permeabilization in cell death. Oncogene. 2008, 27: 6434-6451. Cheung N.S., Koh C.H., Bay B.H., Qi R.Z., Choy M.S., Li Q.T., Wong K.P. and Whiteman M. Chronic exposure to U18666A induces apoptosis in cultured murine cortical neurons. Biochem. Biophys. Res. Commun. 2004, 315:408-417. Chwieralski C., Welte T. and Buhling F. Cathepsin-regulated apoptosis. Apoptosis 2006, 11:143-149. Liao G., Yao Y., Liu J., Yu Z., Cheung S., Xie A., Liang X. and Bi X. Cholesterol accumulation is associated with lysosomal dysfunction and autophagic stress in Npc1-/- mouse brain. Am. J. Pathol. 2007, 171:962-975. This work was supported by: U18666A treatment increases the release of cathepsin D from neurons as detected in the conditioned media of the hippocampal neurons (A and B). Exposure of the hippocampal neurons to conditioned media from U18666A-treated neurons induces loss of viability (C). This toxicity is prevented following removal of cathepsin D using cathepsin D antibody tagged beads (D). Exogenous cathepsin D is toxic to primary hippocampal neurons as detected by MTT (E) and EthD-1 (F and G) assays.