Download

1 / 22
220 likes | 398 Views
Ground state and the glass transition of the RNA secondary structure. Tony Hui and Lei-Han Tang Department of Physics, Hong Kong Baptist University. RNA folding: specific versus nonspecific pairing Ground state and finite temperature properties Logarithmic energy scale

E N D
Ground state and the glass transition of theRNA secondary structure Tony Hui and Lei-Han Tang Department of Physics, Hong Kong Baptist University RNA folding: specific versus nonspecific pairing Ground state and finite temperature properties Logarithmic energy scale Distribution of pairing distances Summary KIAS July 2006
Conformational Characteristics of Biopolymers Complementary Partially complementary sequence specific Increasing complexity and designability Can equilibrium statistical mechanics be of help in understanding bio-specificity? KIAS July 2006
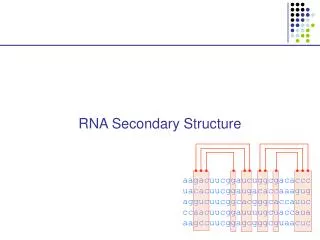
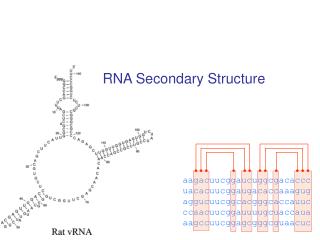
RNA Secondary Structures RNA: single strand molecule of four different nucleotides. Secondary structure: self-matching of the bases. R. Bundschuh and U. Gerland Eur. Phys. J E 19, 319 (2006) Iterative computation of partition function for a finite chain N3 algorithm pairing energy KIAS July 2006
The phase diagramBundschuh and Hwa, PRL 83, 1479 (1999); PRE 65, 031903 (2002). for base pairing Tg Low T: sequence specific pairing High T: nonspecific pairing Michael Laässig and Kay Jörg Wiese, PRL 96, 228101 (2006) KIAS July 2006
Analogy with the directed polymer problem • Transfer matrix power-law algorithm • Ground state scaling properties But what are the values of the exponents for RNA? • Finite temperature transition • Role of disorder distribution KIAS July 2006
Pairing energy • Allowing only Watson and Crick pairing A-U and G-C, but no cooperativity • Extensive g.s. entropy for a typical random sequence • (Higgs, PRL 76, 704 (1996); Pagnani et al. PRL 84, 2026 (2000).) • More realistic energy model (as in Zuker’s Mfold) with stacking energies etc. • force pairing to be at least several nucleotides long (a stem), matching of “words” • Effective model: after coarse graining, we may assume to be independently distributed. A convenient distribution is Adequate for random sequences KIAS July 2006
Pinching (free) energy Bundschuh and Hwa, PRL 83, 1479 (1999); PRE 65, 031903 (2002). (a) Random fluctuations of bond energies largely cancel out probing the effect of a perturbation on large scale. (b) Above the glass transition, (c) A different behavior is expected below KIAS July 2006
Simulation results KIAS July 2006
N = 2 1024 KIAS July 2006
Finite temperatures KIAS July 2006
One or two energy scales on each length scale? Pairing of bases at the end of a sequence is limited Suppose the energy cost due to finite size is proportional to ln N. On each scale, only one such cost is warranted. To insert a break in the middle of the chain, bases close to the mid-point are affected. Hence the energy cost is equal to the sum of costs upto scale N, i.e., KIAS July 2006
Why logarithm? Minimum of N realizations of the pairing energies (energy gain) When pairing with base N+1 splits the chain into two parts. For the pairing to be favorable, we need But this implies pairing will always occur at short distances, in which case the power-law growth is false. KIAS July 2006
Power-law pairing energies Minimum of N2 realizations of the pairing energies Mean and width scale in the same way KIAS July 2006
The surprise Scaled distribution of the site who pairs with the end site Power-law tail at =2 Distribution of pairing distance exponential tail KIAS July 2006
Summary • RNA secondary structure an interesting topic in statistical mechanics, with properties similar to the directed polymer problem. It has a low temperature phase with sequence specific pairing. • Ground state energy of a finite chain contains a log-squared finite size term. • The log-square term persists up to the glass transition, with its coefficient vanishing as the square of the distance to the transition. • Since the energy cost for “remodeling” the pairing pattern grows logarithmically with chain length, two inserted sequences with particularly good matching can easily pair each other, at least under equilibrium conditions. This observation may be of some importance for rRNA’s. • Distribution of the pairing distance assumes a power law with an exponent 4/3, quite independent of the pairing energy distribution. • Analytical treatments? (cf recent attempt by M. Lassig and K. Wiese.) KIAS July 2006
Thank you! KIAS July 2006
RNA World mRNA floppy, ~1000 nt tRNA adaptor, 75-95 nt rRNA Structural, recognition, catalytic, 150-4000 nt snRNA Various functions, e.g., RNA splicing KIAS July 2006
The RNA folding problem primary GCGGAUUUAGCUCAGDDGGGAGAGCGCCAGACUGAAYACUGGAGGUCUGUGT CGAUCCACAGAAUUCGCACCA secondary base pairing 3D structure with tertiary contacts Information flow (from sequence to structure) is hierarchical and sequential. How RNA folds? Tinoco and Bustamante, JMB 293, 271 (1999) KIAS July 2006
Emin E The random energy model B. Derrida, Phys. Rev. B 24, 2613 (1981) N energy levels drawn independently from a probability distribution function (E) glass transition: switching of the dominant term at annealed average typical rare Thermodynamic limit: Engineer the DOS KIAS July 2006
Zuker’s mfold assigning thermodynamic parameters to basic secondary structural components (>1000!) # stack_energies /* CG GC GU UG AU UA -200 -290 -190 -120 -170 -180 -290 -340 -210 -140 -210 -230 -190 -210 150 -40 -100 -110 -120 -140 -40 -20 -50 -80 -170 -210 -100 -50 -90 -90 -180 -230 -110 -80 -90 -110 Task: Identify the ground state configuration among all possible pairing patterns. KIAS July 2006