Download

1 / 70
720 likes | 1.12k Views
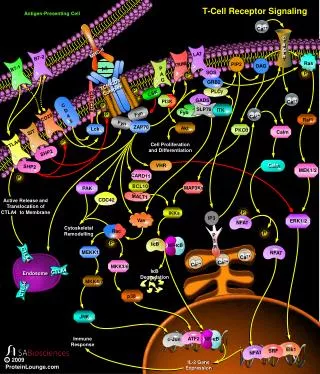
T-cell receptor signalling Toll-like receptor signalling . SIGNAL TRASDUCTION determines CELL RESPONSE. ?. Adaptive Immunity. Innate Immunity. Signal Transduction in Lymphocytes. Cytokine Receptor Signaling Pathways Chemokine Receptor Signaling Pathways

E N D
T-cell receptor signalling • Toll-like receptor signalling
Adaptive Immunity Innate Immunity Signal Transduction in Lymphocytes • Cytokine Receptor Signaling Pathways • Chemokine Receptor Signaling Pathways • Antigen-mediated Signaling Pathways • TCR Signaling • BCR Signaling • NKR Signaling • TLR Signaling • Apoptotic Pathways • Extrinsic stimuli- Fas-FasL, TNFR • Intrinsic stimuli- Stress-DNA damage, UV, growth factor starvation
How Immunity Evolved • The immune system evolved under selective pressure imposed by infectious microorganisms. • Multicellular organisms have developed defense mechanisms that are triggered by infection and protect the host by destroying the invading microbes and neutralizing their virulence factors.
innate dozens germline encoded adaptive millions somatic gene rearrangement How the Immune SystemRecognizes Pathogens immune system RESPONSE: RECEPTORS: DIVERSITY:
Strategies of the Innate Immunity • Recogniction of “microbical non-self”: PAMPS and PPR • Recogniction of “missing self” this concept relies on the detection of “markers on normal self” and was proposed to explain why NK cells preferrentially killed cells that express few or no MHC class I proteins on the cell surface. • Recogniction of “induced or altered self” this third strategy is based on the detection of markers of abnormal self that are induced upon infection (viral infection) and cellular trasformtion. Markers of abnormal self tag the affected cells for elimination by the immune system. Medzhitov and Janeway, Science (2002) vol. 296: 298-300
Recogniction of microbial non-self • Not only is it difficult to recognize pathogens because of their enormous variability and molecular heterogeneity, but microorganisms have an extremely high mutation rate • Good targets for recognition by the innate immune system: 1) are shared by a large number of pathogens and are therefore patterns, and not simply molecular structures 2) must be conserved products of microbial metabolism that do not undergo antigenic variation, and are essential to the survival of the organism 3) structures must the absolutely distinct from self antigens These structures are called Pathogen-Associated Molecular Patterns (PAMPs)
Pattern Recognition Receptors (PRRs) • The principal functions of pattern recognition receptors that recognize PAMPs include opsonization, activation of complement and coagulation cascades, phagocytosis, activation of proinflammatory signaling pathways and induction of apoptosis • In addition, PRRs are a link between innate and adaptive immunity: uptake of the pathogen expressing the PAMPs by APCs can lead to the induction of MHC-peptide molecules as well as costimulatory molecules in order to activate the adaptive immune response • The main difference between PRRs and clonally distributed receptors on B-cells and T-cells is that they are germline-encoded • Pattern recognition receptors can be expressed on the cell surface, in intracellular compartments, or secreted into the bloodstream and tissue fluids
Enter the Toll-like receptor family…. • They are PRRs expressed on the cell surface • Important for the detetion of microrganisms by the innate immune system and triggering of inflammatory response • Play a role in activating the adaptive immune response
Immunity at theHost/Environment Interface innate adaptive gram- bacteria gram+ bacteria yeasts viruses parasites MØ neutrophils APC T cell patterns pattern recognition receptors surface receptors effector molecules counterreceptors
Up-take degli Ag Superficie epiteliale Migrazione Maturazione DC immatura Ag Precursore Presentazione degli Ag Citochine Selezione dei linfociti EO MF NK DC matura Cellule T CTL, helper e regolatorie Organi linfoidi Antigen presenting cells • Dendritic cells • Macrophages • B cells
Enter the Toll Receptor Family… • The Toll protein controls embryonic development in Drosophila, and is required for antifungal immune responses in adult flies. • Toll is a type I transmembrane receptor. The extra-cellular domain contains leucine-rich repeats. The cytoplasmic domain is similar to that of IL-1 receptor. • Ten human homologues of Toll (Toll-like receptors, TLRs) have been identified. • The signaling pathway of the TLR family leads to activation of NF-kB through an adaptor protein, MyD88, and IL-1R-associated kinase (IRAK).
TLRs and Their Ligands lipoproteins (di-/triacylated) lipoarabinomannan LPS (leptospira) LPS (P. gengivalis) PGN (Gram +) zymosan (yeast) GPI anchor (T. cruzi) nucleotides nucleotide analogs LPS (Gram -) taxol (plants) F protein (RSV) Hsp60 (host) fibronectin (host) anti-viral compounds unmethylated CpG DNA flagellin ds RNA MD-2 TLR4 TLR2TLR5TLR9 TLR3 TLR7 TLR6TLR1
The Localization of TLRs TLR3 TLR2 TLR4 Nod2 GP96 endoplasmic reticulum intestinal lumen TLR9 TLR7 TLR5 endosomal compartment
Structure TLRs are type I transmembrane proteins: • Extracellular domain contains leucine-rich repeats • Cytoplasmic Toll/IL-1 receptor (TIR) domain
Signal transduction TLR signaling pathways differ from one another and elicit different biological responses. 2 pathways: • MyD88 – dependent pathway:used by all TLRs • MyD88 – independent pathway:TLR4 and TLR3
MyD88-dependent signaling pathway • MyD88: • adaptor molecule that associates with the TIR domain to TLR TIR domain • recruits the kinase IRAK to the TLR after stimulation of TLR by ligands • IRAK: • Ser/thr kinase • becomes phosphorylated during recruitment to the receptor complex • associates with TRAF6 after phosphorylation • TRAF6: • Induces activation of TAK1 • TAK1: • Activates via NIK and IKK NF-kB Akira, Curr.Op.Immunol. 15: 5, 2003
Focus onTLR4 TLR4 ligand LPS binding complex
Non self ligands LPS ( Gram-negative bacteria) Taxol (plants) Viable Mycobacterium tubercolosis bacilli RSV fusion protein Envelope proteins from mouse mammary tumor virus or from murine leukemia virus Endogenous ligands Mouse -defensin HSP60 (Clamidia and human) HSP70 Fibronectin domain A Satureted fatty acid Minimally modifued LDL TLR4 LIGAND
LPS recogniction complex • LBP, a 60 kDa serum glycoprotein that binds to the lipid A moiety of LPS, is a lipid transferase catalyzing LPS transfer to outer membrane to CD14 • CD14, a glycoprotein expressed on myelomonocytic cells. Mice with a target deletion of the gene encoding CD14 are hyporesponsive to LPS. CD14 seems to have a role in amplifyng LPS responses. • TLR4 • MD-2, is an indispensable molecule for LPS responses. A variety of in vitro studies demostrated that LPS hyporesponsiveness in cells expressing TLR4 alone or TLR4 with mutant MD-2 was rescued bt the transfection of MD-2 cDNA or soluble MD-2 protein.
Four adaptors for TLR4 MyD88 and TIRAP take part in TLR4-mediated proinflammatory cytokine production and B cell activation, but not in induction of the gene encoding IFN- and IFN-inducible genes, the so-called MyD88-independent pathway17, 18, 21, 22. TRIF is essential in the MyD88-independent pathway shared by TLR3 and TLR4 (refs. 25,26). TRAM specifically participates in the MyD88-independent pathway of TLR4, but not TLR3, signaling.
FOCUS on TLR9 • TLR9 ligand: CpG DNA: Bacterial DNA triggers cells of the innate immune system to proliferate and become functionally active. This immune stimulation is mediated by unmethylated CpG motifs present with high frequency in bacterial DNA, but rare in mammalian DNA. The responsiveness to CpG DNA is completely lost in MyD88 KO and, obviously, in TLR9 KO mice. Also in humans, CpG recogniction is mediated by TLR9. • TLR9 subcellular localization: it is expressed on endosomial vescicles • TLR9 signaling pathway
Scheme of subcellular localization of CpG ODN and TLR9 Under physiologic conditions, TLR9 molecules are located below the plasma membrane, presumably in vesicles. Cy3-labeled CpG ODN associate with the cell surface and are transported to these vesicles within 10 min. By 2 h the size and number of CpG ODN-containing endosomes increases significanty, and they relocate from just below the plasma membrane to intracellular regions. In some cases, the CpG ODN and TLR9 molecules co-localize within the same vesicles, recruit MyD88, and initiate the signal transduction process. Takeshita et al. Sem.in Imm. 16 (2004) 17-22
APC-T cell adhesion SIGNAL 2 CO-STIMULATION SIGNAL 1 MHC:pep - TCR Cytokine triggers Receptor IL-2 IL-2R G0 Cytokine Promoter G1 S NUCLEUS IL-2 M G2 Cytokine Production PROLIFERATION DIFFERATION Attivazione delle cellule T
Initial contactStabilization Naive T:DC unique Janeway 5th Edn
All steps Ag non-specific. All common, transient Stability comes from?
Ag alone? Costimulation alone? Ag plus costimulation?
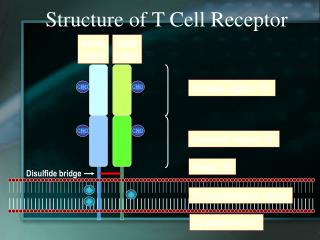
Il complesso del TCR Il TCR è una glicoproteina eterodimerica costituita, nella maggior parte dei linfociti T circolanti, da due diverse catene chiamate a e b. 3 domini: La porzione extracellulare di ogni catena è formata da una regione COSTANTE (C) e da una regione VARIABILE (V) e da una corta regione CERNIERA (HINGE) che contiene i residui di cisteina tra cui si realizza il ponte disolfuro intercatena Un dominio transmembrana, dove sono presenti AA carichi positivamente che interagiscono con le altre molecole di membrana Un corto dominio intracitoplasmaticco rispettivamente di 9 e 12 AA per la catena a e b. Le code intracellulari delle catene a e b sono troppo corte per riuscire a trasdurre il segnale proveniente dall’ingaggio del TCR. Questa funzione viene svolta da un complesso multiproteico di membrana detto CD3 stabilmente associato al TCR. Il complesso CD3 è costituito da tre proteine, omologhe tra loro, dette g,d e e che si associano a formare degli eterodimeri g:e ee:d. Il complesso CD3 è costituito da altre due catene chiamate z e h, le quali si associano tra loro a formare l’omodimero z-z (80% dei casi) o l’eterodimero z-h.
Il complesso del TCR (2): Sequenze ITAM(immunoreceptor-based tyrosine activation-motif) La porzione citoplasmatica di ciascun componente del complesso CD3 e la catena z del TCR contengono un motivo unico detto ITAM (immunoreceptor-based activation motif) costituito dalla sequenza consenso: YxxL/V X(6-8) YxxL/V Le sequenze ITAM sono necessarie e sufficienti per l’attivazione indotto dall’ingaggio del TCR. I residui di Tyr all’interno delle sequenze ITAM vengono rapidamente fosforilate in seguito all’ingaggio del TCR e forniscono dei siti di attacco per il reclutamento in membrana per proteine contenenti motivi SH2. IL PRIMO SEGNALE INTRACELLULARE CHE INDICA CHE I LINFOCITI HANNO RICONOSCIUTO IL LORO ANTIGENE SPECIFICO è LA FOSFORILAZIONE DELLE TYR NELLE ITAM
TCR CD8 Va a Vb b Ca Cb z z/h e g d -S-S- e -S-S- -S-S- p56lck p59fyn Regulation of Src Tyr-Kinase family Phosphorylation of activating tyrosine Regione unica Dominio SH3 Dominio SH2 Dominio chinasico CD45 Csk Tyr attivante Tyr inibitoria Phosphorylation of inhibitory tyrosine residue leads to a conformational change that blocks the active site TCR molecular signalling CD4 CD8 CD3 CD3 -S-S- P P P p56lck C x x C C x C Lck CD8 CD4
CD8 a b -S-S- LcK TCR molecular signalling TCR CD4 Va Vb CD3 CD3 Ca Cb z z/h e g d e -S-S- -S-S- LcK Fyn ON ZAP70 Una volta che le ITAM delle code citoplasmatiche del recettore sono state fosforilate, esse reclutano in membrana la protein-tyr chinasi ZAP70 che viene attivata in seguito a fosforilazione da parte della chinasi LcK Tyrosin Kinase : ZAP/SYK family Y SH2 SH2 Kinase
TCR molecular signalling TCR Va Vb LAT CD3 CD3 Ca Cb z z z/h z/h e g d e e -S-S- -S-S- -S-S- p59fyn PLCg ZAP70 LAT (linker for activation of T cells) è un cosidetto ADATTATORE: contiene una corta sequenza extracellulare, un dominio transmembrana e un ampio dominio citoplasmatico contenente 9 residui di tyr altamente conservati. È associata alla faccia interna della membrana plasmatica mediante residui di cisteina che sono “palmitoilati”. L’associazione di LAT è essenziale per la propagazione del segnale: l’attivazione di alcuni fattori trascrizionali e la produzione di IL-2 non si verifica in cellule T che mancano di LAT.
Altered peptide ligands: APL I peptidi antagonisti inducono una sub-ottimale fosforilazione delle subunita z del CD3 con conseguente assenza di richiamo di ZAP 70 Questo può portare ad anergia delle cellule T
Attivazione di NFAT [Ca2+] Ca2+ RE [Ca2+] LAT CD3 z z z/h z/h e -S-S- -S-S- PIP2 DAG ICRAC IP3 PLCg ZAP70 CALCINEURINA NFAT ON NFAT OFF NFAT
Attivazione della PKC-q TCR complex PLC-g DAG PKC-q IP3 Lck, Fyn, ZAP70 LAT IKKg SEK 1 IKKb IKKa JNK • Incremento [Ca] • intracellulare • Attivazione calcineurina • ON NFAT family IkB DEGRADAZIONE PROTEOSOMA 26S NFkB Attivazione AP1 NUCLEO
La costimolazione amplifica il segnalling del TCR e stabilizza il messaggero per l’IL-2
La costimolazione amplifica il segnalling del TCR e stabilizza il messaggero per l’IL-2
E’ necessario che almeno tre TCR siano attivati sulla sulla superficie cellulare per avere un segnale all’interno della cellula
In seguito ad attivazione la cellule T down-regola il proprio TCR Nel caso in cui una cellula T esprima due TCR, solo il TCR interessato nel legame con il complesso MHC+pep è downregolato Un singolo complesso MHC+peptide puo serialmente attivare molteplici TCR: serial triggering
Costimulation Multiple families of costimulatory molecules APC T cell B7 : B7.1 and B7.2 (CD80, CD86) binds CD28 on T cell (restricted to lymphoid tissues, primarily “activn of naïve T”?) B7-H2, ICOS B7-H3 ??? (both seen widely expressed on non-lymphoid tissues, ie epithelium, primarily interacting with effectors? Not affected by CTLA4) B7 H3 selectively downregulates Th1 activity, favoring Th2: 2003 OX40 CD4 OX40L 4-1BB CD8 4-1BBL (mount normal CD4 or CD8 primary responses, but poorer secondary responses.)
Multiple families of costimulatory molecules APC T cell B7 : B7.1 and B7.2 (CD80, CD86) binds CD28 on T cell Disruption leads to very weak immune responses ~10% of normal. Thus “the” major co-stimulatory family for initiating T cell immunity B7-H2, ICOS B7-H3 ??? Ox40 (CD4) OX40L 4-1BB (CD8) 4-1BBL Disruption results in subtle effects, such as attenuated or different secondary responses; different types of response (Th1 vs Th2 bias). Bottom line: Multiple families of costimulatory molecules are found to direct nuances of immune responses: sustaining or differentiating immune responses. Nuance: IgM vs IgE to parasite, so not trivial
SINAPSI IMMUNOLOGICA Sull’APC, quando il TCR Riconosce il complesso MHC-peptide, numerose proteine di membrana e intracellulari vengono mobilizzate rapidamente. La regione di contatto fisico tra APC e cellula T È chiamata SINAPSI IMMUNOLOGICA o CLUSTER di ATTIVA- ZIONE SOPRAMOLE- COLARE (SMAC). TCR, CD3, catene , i corecettori CD4 o CD8, e il CD28 sono mobilizzati al centro della sinapsi. Le integrine restano invece alla sua periferia. cSMAC= central Supramolecular Cluster pSMAC= periferal Supramolecular Cluster
Analisi in “imaging al microscopio confocale ” della formazione della sinapsi immunologica Una cell.T specifica per un determinato complesso MHC-peptide lega un’APC. CD3 (verde) si localizza sullo cSMAC. L’integrina LFA-1 (rosso) invece si localizza sullo pSMAC. CD3 è al centro e LFA-1 è alla periferia della sinapsi immunologica.
B7.1 e B7.2: omodimeri, membri della superfamiglia delle Ig se trasfettate in fibroblasti conferiscono capacità costimolatoria CD28. Glicoproteina espressa costitutivamente sulla superficie dell’80% delle cellule T umane (tutte le CD4 e il 50% delle CD8) e sul 100% delle cellule T murine. e’ upregolato in seguito ad attivazione.