DNA STRUCTURE
310 likes | 582 Views
DNA STRUCTURE. STRUCTURE, FORCES AND TOPOLOGY. DNA GEOMETRY. A POLYMER OF DEOXYRIBONUCLEOTIDES DOUBLE-STRANDED INDIVIDUAL deoxyNUCLEOSIDE TRIPHOSPHATES ARE COUPLED BY PHOSPHODIESTER BONDS ESTERIFICATION LINK 3’ CARBON OF ONE RIBOSE WITH 5’ C OF ANOTHER TERMINAL ENDS : 5’ AND 3’

DNA STRUCTURE
E N D
Presentation Transcript
DNA STRUCTURE STRUCTURE, FORCES AND TOPOLOGY
DNA GEOMETRY • A POLYMER OF DEOXYRIBONUCLEOTIDES • DOUBLE-STRANDED • INDIVIDUAL deoxyNUCLEOSIDE TRIPHOSPHATES ARE COUPLED BY PHOSPHODIESTER BONDS • ESTERIFICATION • LINK 3’ CARBON OF ONE RIBOSE WITH 5’ C OF ANOTHER • TERMINAL ENDS : 5’ AND 3’ • A “DOUBLE HELICAL” STRUCTURE • COMMON AXIS FOR BOTH HELICES • “HANDEDNESS” OF HELICES • ANTIPARALLEL RELATIONSHIP BETWEEN 2 DNA STRANDS
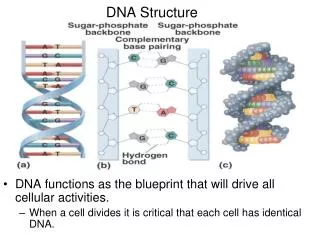
DNA GEOMETRY • PERIPHERY OF DNA • SUGAR-PHOSPHATE CHAINS • CORE OF DNA • BASES ARE STACKED IN PARALLEL FASHION • CHARGAFF’S RULES • A = T • G = C • “COMPLEMENTARY” BASE-PAIRING
TAUTOMERIC FORMS OF BASES • TWO POSSIBILITIES • KETO (LACTAM) • ENOL (LACTIM) • PROTON SHIFTS BETWEEN TWO FORMS • IMPORTANT IN ORDER TO SPECIFY HYDROGEN BONDING RELATIONSHIPS • THE KETO FORM PREDOMINATES
MAJOR AND MINOR GROOVES • MINOR • EXPOSES EDGE FROM WHICH C1’ ATOMS EXTEND • MAJOR • EXPOSES OPPOSITE EDGE OF BASE PAIR • THE PATTERN OF H-BOND POSSIBILITIES IS MORE SPECIFIC AND MORE DISCRIMINATING IN THE MAJOR GROOVE • STUDY QUESTION: LOCATE ALL OF THE POSSIBILITIES FOR H-BONDING IN THE MAJOR AND MINOR GROOVES FOR THE 4 POSSIBLE BASE-PAIRS
STRUCTURE OF THE DOUBLE HELIX • THREE MAJOR FORMS • B-DNA • A-DNA • Z-DNA • B-DNA IS BIOLOGICALLY THE MOST COMMON • RIGHT-HANDED (20 ANGSTROM (A) DIAMETER) • COMPLEMENTARY BASE-PAIRING (WATSON-CRICK) • A-T • G-C • EACH BASE PAIR HAS ~ THE SAME WIDTH • 10.85 A FROM C1’ TO C1’ • A-T AND G-C PAIRS ARE INTERCHANGEABLE • “PSEUDO-DYAD” AXIS OF SYMMETRY
GEOMETRY OF B-DNA • IDEAL B-DNA HAS 10 BASE PAIRS PER TURN • BASE THICKNESS • AROMATIC RINGS WITH 3.4 A THICKNESS TO RINGS • PITCH = 10 X 3.4 = 34 A PER COMPLETE TURN • AXIS PASSES THROUGH MIDDLE OF EACH BP • MINOR GROOVE IS NARROW • MAJOR GROOVE IS WIDE • IN CLASS EXERCISE: EXPLORE THE STRUCTURE OF B-DNA. PAY SPECIAL ATTENTION TO THE MAJOR, MINOR GROOVES
A-DNA • RIGHT-HANDED HELIX • WIDER AND FLATTER THAN B-DNA • 11.6 BP PER TURN • PITCH OF 34 A • AN AXIAL HOLE • BASE PLANES ARE TILTED 20 DEGREES WITH RESPECT TO HELICAL AXIS • HELIX AXIS PASSES “ABOVE” MAJOR GROOVE • DEEP MAJOR AND SHALLOW MINOR GROOVE • OBSERVED UNDER DEHYDRATING CONDITIONS
A-DNA • WHEN RELATIVE HUMIDITY IS ~ 75% • B-DNA A-DNA (REVERSIBLE) • MOST SELF-COMPLEMENTARY OLIGONUCLEO- TIDES OF < 10 bp CRYSTALLIZE IN A-DNA CONF. • A-DNA HAS BEEN OBSERVED IN 2 CONTEXTS: • AT ACTIVE SITE OF DNA POLYMERASE (~ 3 bp ) • GRAM (+) BACTERIA UNDERGOING SPORULATION • SASPs INDUCE B-DNA TO A-DNA • RESISTANT TO UV-INDUCED DAMAGE • CROSS-LINKING OF PYRIMIDINE BASES
Z-DNA • A LEFT-HANDED HELIX • SEEN IN CONDITIONS OF HIGH SALT CONCENTRATIONS • REDUCES REPULSIONS BETWEEN CLOSEST PHOSPHATE GROUPS ON OPPOSITE STRANDS (8 A VS 12 A IN B-DNA) • IN COMPLEMENTARY POLYNUCLEOTIDES WITH ALTERNATING PURINES AND PYRIMIDINES • POLY d(GC) · POLY d(GC) • POLY d(AC) POLY d(GT) • MIGHT ALSO BE SEEN IN DNA SEGMENTS WITH ABOVE CHARACTERISTICS
Z-DNA • 12 W-C BASE PAIRS PER TURN • A PITCH OF 44 DEGREES • A DEEP MINOR GROOVE • NO DISCERNIBLE MAJOR GROOVE • REVERSIBLE CHANGE FROM B-DNA TO Z-DNA IN LOCALIZED REGIONS MAY ACT AS A “SWITCH” TO REGULATE GENE EXPRESSION • ? TRANSIENT FORMATION BEHIND ACTIVELY TRAN- SCRIBING RNA POLYMERASE
STRUCTURAL VARIANTS OF DNA • DEPEND UPON: • SOLVENT COMPOSITION • WATER • IONS • BASE COMPOSITION • IN-CLASS QUESTION: WHAT FORM OF DNA WOULD YOU EXPECT TO SEE IN DESSICATED BRINE SHRIMP EGGS? WHY?
RNA • UNLIKE DNA, RNA IS SYNTHESIZED AS A SINGLE STRAND • THERE ARE DOUBLE-STRANDED RNA STRUCTURES • RNA CAN FOLD BACK ON ITSELF • DEPENDS ON BASE SEQUENCE • GIVES STEM (DOUBLE-STRAND) AND LOOP (SINGLE-STRAND STRUCTURES) • DS RNA HAS AN A-LIKE CONFORMATION • STERIC CLASHES BETWEEN 2’-OH GROUPS PREVENT THE B-LIKE CONFORMATION
HYBRID DNA-RNA STRUCTURES • THESE ASSUME THE A-LIKE CONFORMATION • USUALLY SHORT SEQUENCES • EXAMPLES: • DNA SYNTHESIS IS INITIATED BY RNA “PRIMERS” • DNA IS THE TEMPLATE FOR TRANSCRIPTION TO RNA
FORCES THAT STABILIZE NUCLEIC ACID STRUCTURES • SUGAR-PHOSPHATE CHAIN CONFORMATIONS • BASE PAIRING • BASE-STACKING,HYDROPHOBIC • IONIC INTERACTIONS
SUGAR-PHOSPHATE CHAIN IS FLEXIBLE TO AN EXTENT • CONFORMATIONAL FLEXIBILITY IS CONSTRAINED BY: • SIX TORSION ANGLES OF SUGAR-PHOSPHATE BACKBONE • TORSION ANGLES AROUND N-GLYCOSIDIC BOND • RIBOSE RING PUCKER
TORSION ANGLES • SIX OF THEM • GREATLY RESTRICTED RANGE OF ALLOWABLE VALUES • STERIC INTERFERENCE BETWEEN RESIDUES IN POLYNUCLEOTIDES • ELECTROSTATIC INTERACTIONS OF PHOS. GROUPS • A SINGLE STRAND OF DNA ASSUMES A RANDOM COIL CONFIGURATION
THE N-GLYCOSIDIC TORSION ANGLE • TWO POSSIBILITIES, STERICALLY • SYN • ANTI • PYRIMIDINES • ONLY ANTI IS ALLOWED • STERIC INTERFERENCE BETWEEN RIBOSE AND THE C2’ SUBSTITUENT OF PYRIMIDINE • PURINES • CAN BE SYN OR ANTI
GLYCOSIDIC TORSION ANGLES IN Z-DNA • ALTERNATING • PYRIMIDINE: ANTI • PURINE: SYN • WHAT HAPPENS WHEN B-DNA SWITCHES TO Z-DNA? • THE PURINE BASES ROTATE AROUND GLYCOSIDIC BOND FROM ANTI TO SYN • THE SUGARS ROTATE IN THE PYRIMIDINES • THIS MAINTAINS THE ANTI CONFORMATIONS
RIBOSE RING PUCKER • THE RING IS NOT FLAT • SUBSTITUENTS ARE ECLIPSED IF FLAT • CROWDING IS RELIEVED BY PUCKERING • TWO POSSIBILITIES FOR EACH OF C2’ OR C3’: • ENDO: OUT-OF-PLANE ATOM ON SAME SIDE OF RING AS C5’ • EXO; DISPLACED TO OPPOSITE SIDE • C2’ ENDO IS MOST COMMON • CAN ALSO SEE C3’-ENDO AND C3’-EXO • LOOK AT RELATIONSHIPS BETWEEN THE PHOSPHATES: • IN C3’ ENDO- THE PHOSPHATES ARE CLOSER THAN IN C2’ ENDO-
RIBOSE RING PUCKER • B-DNA HAS THE C2’-ENDO-FORM • A-DNA IS C3’-ENDO • Z-DNA • PURINES ARE ALL C3’-ENDO • PYRIMIDINES ARE ALL C2’-ENDO • CONCLUSION: THE RIBOSE PUCKER GOVERNS RELATIVE ORIENTATIONS OF PHOSPHATE GROUPS TO EACH SUGAR RESIDUE
IONIC INTERACTIONS • THE DOUBLE HELIX IS ANIONIC • MULTIPLE PHOSPHATE GROUPS • DOUBLE-STRANDED DNA HAS HIGHER ANIONIC CHARGE DENSITY THAT SS-DNA • THERE IS AN EQUILIBRIUM BETWEEN SS-DNA AND DS-DNA IN AQUEOUS SOLUTION: • DS-DNA == SS-DNA • QUESTION: WHAT HAPPENS TO THE Tm OF DS-DNA AS [CATION] INCREASES? WHY?
IONIC INTERACTIONS • DIVALENT CATIONS ARE GOOD SHIELDING AGENTS • MONOVALENT CATIONS INTERACT NON-SPECIFICALLY • FOR EXAMPLE, IN AFFECTING Tm • DIVALENT INTERACT SPECIFICALLY • BIND TO PHOSPHATE GROUPS • MAGNESIUM (2+) ION • STABILIZES DNA AND RNA STRUCTURES • ENZYMES THAT ARE INVOLVED IN RXNS’ WITH NUCLEIC ACID USUALLY REQUIRE Mg(2+) IONS FOR ACTIVITY
BASE STACKING • PARTIAL OVERLAP OF PURINE AND PYRIMIDINE BASES • IN SOLID-STATE (CRYSTAL) • VANDERWAALS FORCES • IN AQUEOUS SOLUTION • MOSTLY HYDROPHOBIC FORCES • ENTHALPICALLY-DRIVEN • ENTROPICALLY-OPPOSED • OPPOSITE TO THAT OF PROTEINS
BASE-PAIRING • WATSON-CRICK GEOMETRY • THE A-T PAIRS USE ADENINE’S N1 AS THE H-BOND ACCEPTOR • HOOGSTEEN GEOMETRY • N7 IS THE ACCEPTOR • SEEN IN CRYSTALS OF MONOMERIC A-T BASE PAIRS • IN DOUBLE HELICES, W-C IS MORE STABLE • ALTHOUGH HOOGSTEIN IS MORE STABLE FOR A-T PAIRS, W-C IS MORE STABLE IN DOUBLE HELICES • CO-CRYSTALLIZED MONOMERIC G-C PAIRS ALWAYS FOLLOW W-C GEOMETRY • THREE H-BONDS
HYDROGEN BONDING • REQUIRED FOR SPECIFICITY OF BASE PAIRING • NOT VERY IMPORTANT IN DNA STABILIZATION • HYDROPHOBIC FORCES ARE THE MOST IMPT.’
THE TOPOLOGY OF DNA • “SUPERCOILING” : DNA’S “TERTIARY STRUCTURE • L = “LINKING NUMBER” • A TOPOLOGIC INVARIANT • THE # OF TIMES ONE DNA STRAND WINDS AROUND THE OTHER • L = T + W • T IS THE “TWIST • THE # OF COMPLETE REVOLUTIONS THAT ONE DNA STRAND MAKES AROUND THE DUPLEX AXIS • W IS THE “WRITHE” • THE # OF TIMES THE DUPLEX AXIS TURNS AROUND THE SUPERHELICAL AXIS
DNA TOPOLOGY • THE TOPOLOGICAL PROPERTIES OF DNA HELP US TO EXPLAIN • DNA COMPACTING IN THE NUCLEUS • UNWINDING OF DNA AT THE REPLICATION FORK • FORMATION AND MAINTENANCE OF THE TRANSCRIPTION BUBBLE • MANAGING THE SUPERCOILING IN THE ADVANCING TRANSCRIPTION BUBBLE
DNA TOPOLOGY • AFTER COMPLETING THE 13 IN-CLASS EXERCISES, TRY TO ANSWER THE FOLLOWING QUESTIONS: • (1) THE HELIX AXIS OF A CLOSED CIRCULAR DUPLEX DNA IS CONSTRAINED TO LIE IN A PLANE. THERE ARE 2340 BASE PAIRS IN THIS PIECE OF DNA AND, WHEN CONSTRAINED TO THE PLANE, THE TWIST IS 212. • DETERMINE “L”, “W” AND “T” FOR THE CONSTRAINED AND UNCONSTRAINED FORM OF THIS DNA. • (2) A CLOSED CIRCULAR DUPLEX DNA HAS A 100 BP SEGMENT OF ALTERNATING C AND G RESIDUES. ON TRANSFER TO A SOLUTION WITH A HIGH SALT CONCENTRATION, THE SEGMENT MAKES A TRANSITION FROM THE B-FORM TO THE Z-FORM. WHAT IS THE ACCOMPANYING CHANGE IN “L”, “W”. AND “T”?