Download
1 / 37
370 likes | 389 Views
This article explores the mapping and dynamics of nucleosome positions in gene regulation, including the role of chromatin remodeling, histone modifications, and heterochromatin formation. It discusses the impact of these processes on gene expression and chromatin structure.

E N D
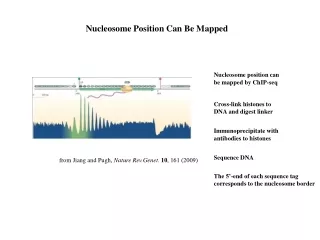
Nucleosome Position Can Be Mapped Nucleosome position can be mapped by ChIP-seq Cross-link histones to DNA and digest linker Immunoprecipitate with antibodies to histones Sequence DNA from Jiang and Pugh, Nature Rev.Genet. 10, 161 (2009) The 5’-end of each sequence tag corresponds to the nucleosome border
Positioning of Nucleosomes at Promoters Nucleosome-free region at the beginning and end of genes Nucleosomes have defined locations near the promoter Nucleosome position is determined by ATP-dependent trans-acting factors from Jiang and Pugh, Nature Rev.Genet. 10, 161 (2009)
Nucleosome Dynamics During Transcription Initiation A chromatin remodeller removes the nucleosome from the AT-rich promoter region Histone chaperones replace H3-H4 with H3.3-H4 and H2A-H2B with H2A.Z-H2B Retention of H2A.Z over the promoter is ensured by its acetylation preventing its exchange by INO80 H2A.Z prevents methylation of promoter DNA From Venkatesh and Workman, Nature Rev.Mol.Cell Biol.16, 178 (2015)
Nucleosome Dynamics During Transcription Elongation From Venkatesh and Workman, Nature Rev.Mol.Cell Biol.16, 178 (2015) Removal of H2A-H2B dimer is sufficient to allow Pol II passage FACT targets ubiquitylated H2A-H2B dimers for removal Nap1 stabilizes hexameric nucleosome
Histone Tails Histones contain flexible termini that extend from the globular structure of the nucleosome from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-31
Modification of Histone Tails Chromatin structure is a source of epigenetic information Posttranslational modifications and histone variants contribute to structural and functional characteristics of chromatin The combination of histone modifications constitutes the histone code The histone code influences chromatin condensation and function and defines actual or potential transcription states from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-31
Histone Modifications Associated with Heterochromatin and Euchromatin from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-33
Histone Acetylation from Tessarz and Kouzarides, Nature Rev.Mol.Cell Biol. 15, 703 (2014) The positive charge of lys residues can form a salt bridge with DNA Histone acetylation removes the positive charge decreasing the binding affinity to DNA and nucleosome stability
Control of Gene Expression by Acetylation Repressor recruits a complex that contains a histone deacetylase Neighboring histones are deacetylated Activator recruits a complex that contains a histone acetyltransferase Neighboring histones are acetylated from Lodish et al., Molecular Cell Biology, 6th ed. Fig 7-38
Effect of Histone H3 K9 Methylation SUV39 methylates K9 Methylated K9 recruits HP1 Heterochromatin formation HP1 binds to SUV39 to propagate methylation Acetylated K9 or phosphorylated S10 inhibits methylation of K9 from Turner, Cell111, 285 (2002)
Effect of Histone H3K4 Methylation Set9 methylates K4 Inhibits association of NuRD remodeling and deacetylase complex Inhibits association of SUV39 H3K4me is associated with active genes from Turner, Cell111, 285 (2002)
Histone Modifications Affect Chromatin Structure H3K4 methylation and H3K9 acetylation are hallmarks of active chromatin H3K27 methylation and H3K9 methylation are hallmarks of silent chromatin from Johnstone and Baylin, Nature Rev.Genet. 11, 806 (2010)
Histone Modifications Define Functional Elements Each histone modification has a unique biological role Histone modifications are interdependent from Zhou et al., Nature Rev.Mol.Cell Biol. 12, 7 (2011)
Two Major States of Chromatin from Dawson, Science355, 1147 (2017) Heterochromatin is more compact than euchromatin and is largely transcriptionally silent DNA and histones are reversibly modified Epigenetic regulators bind chromatin modifications to facilitate DNA templated processes
Properties of Heterochromatin Darkly stained and condensed Transcriptionally silent and silences adjacent genes Present at centromeres and telomeres HP1 interacts with H3 only when K9 is methylated Repressive structure can be propagated Euchromatic gene placed in heterochromatin is repressed from Lodish et al., Molecular Cell Biology, 6th ed. Fig 6-33
Variations in Heterochromatin Structure Constitutive heterochromatin is present in centromeres and repeat sequences, and contains H3K9me3 Facultatitive heterochromatin is dynamic during development, and contains H3K27me3 Sonication-resistant heterochromatin is “poised” for gene expression despite the presence of H3K27me3 from Tchasovnikarova and Kingston, Molecular Cell 69, 5 (2018)
Epigenome Modification and Interpretation Writers catalyze posttranslational modifications on DNA or proteins Erasers remove posttranslational modifications and DNA methylation Readers interpret the modifications and alter chromatin structure from Helin and Dhanak, Nature502, 480 (2013)
Structure and Propagation of Heterochromatin from Reinberg and Vales, Science 361, 33 (2018) Heterochromatin spreads by reader-writer coupling In facultative heterochromatin, EED reads H3K27me3. Binding stimulates EZH2 writer activity In constitutive heterochromatin, HP1 reads H3K9me3. Binding stimulates SUV39H1 writer activity
Heterochromatin Assembly in S. pombe Requires Transcription Repeat sequences are transcribed and converted to dsRNA dsRNA is processed to siRNA siRNA is loaded onto the RITS complex and targeted to nascent transcripts RITS associates with Clr4 which methylates H3K9 from Wang et al., Trends Genet.32, 284 (2016)
Phase Separation Contents of cells can separate into droplets to form membrane-less structures Phase separation clusters components together to speed up reactions or sequester unwanted molecules from Dolgin, Nature555, 300 (2018)
Examples of Membraneless Organelles from Boeynaems et al., Trends Cell Biol. 28, 420 (2018) Membraneless organelles have different physical properties (solute, liquid, gel, solid) Formation of membraneless organelles by phase transition is mediated by intrinsically disordered protein and often contain RNA Proteins that are involved in neurodegenerative disorders are components of membraneless organelles
Heterochromatin Compaction by Phase Separation HP1 domains undergo phase separation and form liquid-like droplets Phase separation in heterochromatin can exclude proteins (transcription factors) from Klosin and Hyman, Nature547, 168 (2017)
Mechanisms of Heterochromatin Barrier Formation Pol III binding to tRNA genes can generate nucleosome depleted regions The Pol II elongation factor, Paf1, promotes nucleosome turnover Transcription units mediate the addition of active histone modifications and H2A.Z Readers can recruit erasers to limit heterochromatin spread The Epe1 eraser is degraded by a Ub ligase which is active in the interior of heterochromatin from Allshire and Madhani, Nature Rev.Mol.Cell.Biol. 19, 229 (2018)
Heterochromatin is Maintained Following DNA Replication from Maison and Almounzi, Nature Rev.Mol.Cell Biol. 5, 296 (2004) Passage of the replication fork releases parental modified nucleosomes Nucleosome binding sites are created by recruitment of CAF1 by PCNA CAF1-bound HP1 recruits Suv39h, Dnmt1, and HDAC Methylated histones provide new HP1 binding sites
Heterochromatin Functions DNA or H3 methylation recruits adaptors such as HP1 Adaptors recruit effectors that are involved in chromosome segregation, gene silencing, transcriptional activation, and histone modification from Grewal and Gia, Nature Rev.Genet.8, 35 (2007)
Epigenetics Heritable changes in gene function that cannot be explained by changes in gene sequences DNA methylation Histone variants and modifications Nucleosome positioning Cells retain a memory of past states Epigenetic information is involved in cell fate determination and stability
Epigenetic Modifications During Development Epigenetically imposed restrictions to plasticity are erased in the germ line Early mammalian development is characterized by progressive restriction of cellular plasticity accompanied by acquisition of epigenetic modifications Epigenetic modifications impose a cellular memory that accompanies and enables stable differentiation
DNA Methylation Methylation at CpG residues correlates with gene repression 5meC is involved in stable epigenetic repression Sites of methylation Inactive X Imprinted loci Transposon-derived sequences CpG islands are CpG-rich regions usually found at promoters Methylation patterns are reproduced at each round of cell division Organisms that lack DNA methylation have cellular memory
Methylated CpG Islands Inhibit Transcription from Portela and Esteller, Nature Biotechnol. 28, 1057 (2010) More than half of human promoters contain CpG islands Promoters are usually unmethylated Methylated DNA recruits methyl-CpG-binding domain proteins which recruit histone modifying and chromatin-remodelling complexes Unmethylated CpG islands recruit Cfp1 which associates with a histone methyltransferase creating H3K4me3
Establishment of DNA Methylation Pattern Most CpGs are unmethylated before implantation RNA pol II recruits H3K4 methyltransferase DNMT3L only binds unmethylated H3K4 and recruits DNA methyltransferases from Cedar and Bergman, Nature Rev.Genet. 10, 295 (2009)
DNA Methylation UHRF1 binds hemimethylated DNA and recruits DNMT1 H3K36me recruits DNMT3B to establish de novo symmetric methylation CTCF and cohesin target DNMT3A for hemimethylation Hemimethylation may play a role in long-range chromatin interactions from Sharif and Koseki, Science359, 1102 (2018)
Propagation of DNA Methylation State Newly synthesized methylated DNA is hemimethylated NP95 binds hemimethylated DNA DNMT1 is a maintenance methyltransferase and binds PCNA NP95 links DNMT1 to hemimethylated DNA from Richly et al., BioEssays32, 669 (2010)
DNA is Demethylated by TET Proteins 5mC is oxidated iteratively by TET 5hmC is reverted to unmodified C by passive dilution during DNA replication Oxidative products are excised by thymine DNA glycosylase and repaired by BER from Kohli and Zhang, Nature502, 472 (2013)
Rett Syndrome Onset of symptoms in females is 6-12 months of age Rett syndrome is a severe neurological disorder Abnormal head growth Decrease in speech function Breathing disturbances Repetitive hand movements Caused by mutation in the MeCP2 gene on the X chromosome Severity of symptoms varies with the proportion of cells expressing the mutant copy of MeCP2
MeCP2 Function and Rett Syndrome from Lyst and Bird, Nature Rev.Genet. 16, 261 (2015) MeCP2 binds methylated DNA, but also binds to unmethylated DNA via regions outside the MBD MeCP2 compacts chromatin structure MeCP2 recruits a corepressor complex that contains a histone deacetylase MeCP2 can also activate gene expression
MeCP2 Regulates Gene Expression in Response to Neural Activity MeCP2 binds methylated DNA and silences target genes such as BDNF and corticotropin-releasing hormone Neural activity triggers MeCP2 phosphorylation and target gene activation from Bienvenu and Chelly, Nature Rev.Genet.7, 415 (2006) Hippocampal neurons grow dendrites with fewer branches when MeCP2 is blocked Increase in mCH sequences after birth is enriched in genes with neuronal functions from Miller, Science314, 1356 (2006)
Many MeCP2-Repressed Genes Encode Proteins That Modulate Neuronal Physiology MeCP2 binds mCA Density of mCA is higher in long genes Frequency of mCA increases with age Long genes are selectively expressed in the brain Length- and mCA-dependent increase in gene expression in MeCP2 mutants Increased binding of MeCP2 at the BDNF locus in adult brains from Gabel et al., Nature522, 89 (2015) from Luo and Ecker, Science348, 1094 (2015)