Nucleic Acids Structures
Nucleic Acids Structures. 1-Discovery of DNA structure. 2- A, B and Z conformations of dsDNA/dsRNA. 3- DNA tertiary Structures Nucleosome , G- quadruplex. 4- Secondary and tertiary Structure of RNA. 5- Principles of DNA Recognition by sequence-specific DNA binding proteins .

Nucleic Acids Structures
E N D
Presentation Transcript
Nucleic Acids Structures 1-Discovery of DNA structure 2- A, B and Z conformations of dsDNA/dsRNA 3- DNA tertiary Structures Nucleosome, G-quadruplex 4- Secondary and tertiary Structure of RNA 5- Principles of DNA Recognition by sequence-specific DNA binding proteins • • Not treated: • DNA topology • DNA Sequencing 6- Principles of Nucleic Acids Denaturation
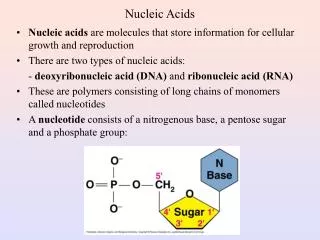
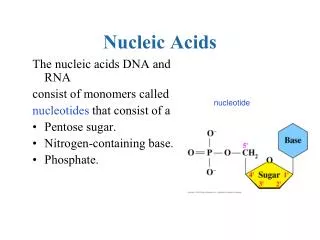
Polymeric Structure Of Nucleic Acids • Links 3’-O of preceding nucleotide to 5’ of next nucleotide ->5’-3’ polarity • 1 negative charge per residue
Rosalind Franklin (1950 or 1951) Chargaff. 1950: “It is, however, noteworthy -whether this is more than accidental, cannot yet be said-that in all deoxypentose nucleic acids examined thus far the molar ratios of total purines to total pyrimidines, and also of adenine to thymine and of guanine to cytosine, were not far from 1”. Watson and Crick (1953) Watson and Crick (1953): “It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material”.
Information that Watson and Crick used to propose the double helix model: 1) R. Franklin DNA fibers X-ray diffraction data 2) bases are in the keto conformation Maltese Cross Indicates an Helical pattern Spacing between Phosphates = 3.4A enol Helical Pitch = 34A keto Bragg’s Law: 2dsinQ = nl used to interpret X-ray diffraction pictures 4) Density measurements: ~2 polymers/helix 3) Chargaff’s rules: (G+C)/(A+T) can vary But (G+A)/(C+T) = G/C = A/T =1 5) C2’ endo sugar pucker conformation
Bragg's Law = repeated atomic features in the crystal or fiber Bragg’s law indicates an inverse relationship between diffraction angle and actual distances between repeated features in crystal/fiber
The original model for DNA structure Watson and Crick (1953) Nature 171, 964-967 • Essential features of the model that • proved correct: • Antiparallel right-handed double helix • 2) Strands are linked by • complementary sets of • donors and acceptor groups • on bases Helical Pitch = 34 A (10 residues/turn) Rise/ residue = 3.4 A
A comparison of the Watson-Crick model (1953) and of the first B-DNA structure solved (1980) The Dickerson Dodecamer X-ray structure (CGCGAATTCGCG) PDB ID: 1BNA Watson-Crick Model
DNA Double Helix Definitions Bases Orientation
Base pairs seen from above the helix (helical projection) Helical Axis Major Groove >180° H O H N N7 N1 H N3 N9 N1 N3 O H N dR dR H Minor Groove <180° Pseudo Dyad Axis
H H N O CH3 N7 A T N1 H N3 N9 N1 N3 O dR dR H O H CH3 N N7 T A N1 H N3 N9 N1 N3 dR O dR dR dR O CH3 H T O H H N N3 O G N7 N7 N1 G C O N3 H N1 N1 H N9 N9 N1 N3 N3 O N H H N dR dR H H H H O O H H N N G G C C N7 N7 N1 N1 H H N3 N3 N9 N9 N1 N1 N3 N3 O O H H N N dR dR dR dR H H Isostericity of Watson-Crick Base Pairs (and non isostericity of non WC base pairs) Example of a G-T non WC base pair
B A vs B DNA H20 A B PDB ID: 1BNA Ethanol A PDB ID = 115D
Sugar Pucker Planar C3’endo C2’endo Exact values need not to be remembered… helical projection Major differences : - A DNA is shorter than B DNA: 1 helix turn is 28.6A vs 34 A for B DNA. This is due to the 3’ endo sugar pucker in A - The Bases of A-DNA are shifted away from the helical axis. This results in a deep major groove and in a shallow minor groove. There is a 6 A hole in a helical projection. A B
Sugar puckering: C2’ endo or C3’ endo dsRNA or A-DNA : C3’ endo B-DNA: C2’ endo Distance between Consecutive Phosphates: 5.9 Å 7 Å
Base tilting in A-DNA B-DNA A-DNA Base pairs are more tilted in A-DNA.
H20 is essential in the transition A <--> B DNA H20 A B A water spine (green dots) has been proposed to exist in the minor groove of B-DNA that would stabilize the B-form This concept is controversial and will not be detailed further
Z-DNA Left handed Helix Occurs in DNA sequences with stretches of consecutive G-C base pairs jagged backbone G nucleotides: Switch C2’endo -> C3’ endo anti -> Syn Requires high salt in vitro PDB ID: 1DCG C nucleotides: No change www.mun.ca/biology/scarr/A_B_Z_DNA.html
Nucleotides flipping and grooves in Z-DNA Z-DNA Major Minor Major Minor B-DNA Note: this simplified diagram only summarizes the conformation changes during the B->Z transition – it does not accurately shows the Z structure
Glycosidic bond Anti /Syn conformations Anti and Syn conformations are defined based on the torsion angle of the glycosidic bond The sequence of atoms chosen to define the torsion angle to define anti/syn conformation is: O4'-C1'-N9-C4 for purines - O4'-C1'-N1-C2 for pyrimidines. Anti C/T: C1’-O4’ and N1-C2 are pointing away from each other Syn A/G: C1’-O4’ and N9-C4 are pointing in same direction Anti A/G: C1’-O4’ and N9-C4 are pointing away from each other 4 4 2 9 9 1 4' 4' 4' 1' 1' 1'
Anti /Syn conformations in pseudo-3D 9 4 4' 4 4' 9 1' 1' Syn- A 4 9 4' 4' 4 9 1' 1' Anti A/G: C1’-O4’ and N9-C4 are pointing away from each other Syn A/G: C1’-O4’ and N9-C4 are pointing in same direction
B Z A ABZ-DNAs B Z Backbone Profiles Helical Projections
Z A B www.mun.ca/biology/scarr/A_B_Z_DNA.html
Why Study DNA Structure ? • Structure and Sequence Recognition by DNA binding proteins • Some non B-DNA structures are biologically relevant - dehydrated living forms - dsRNA is A form (see PDB: 2KYD) - DNA/RNA duplex (replication, transcription) is A form - Z-DNA might be associated with promoter elements, regulatory sequences • There are conformations other than A/B/Z e.g.: conformations intermediate between A and B Also Tertiary conformation of DNA
Double-Stranded DNA is wrapped around nucleosomes in eukaryotic cells Binding of histones to DNA through electrostatic interactions: Histones are + charged, DNA is - charged Consequences for: DNA Replication, DNA Repair Transcription http://www.bio.miami.edu/dana/104/nucleosome.jpg
Double-Stranded DNA is wrapped around nucleosomes in eukaryotic cells PDB ID: 1AOI http://www.chem.ucsb.edu/~molvisual/dna_biochem.html
Example of intrinsic DNA Tertiary Structure G-quadruplex structures in telomeric DNA: case of (T2G4) repeats Na+ PDB ID: 156D
Secondary and Tertiary Structure of RNA Single strandedness nature of RNA makes it able to “fold” on itself and base-pair with complementary segments within the same molecule
Secondary and Tertiary Structure of transfer RNA (tRNA) PDB ID = 3TRA
Secondary and Tertiary Structure of RNA: See other examples in the RNA Processing and Translation Chapters Secondary Structure of the M1 RNA, a component of RNase P (see RNA processing chapter) Two major observations: 1-Abundance of G:U base pairs 2-Pseudoknot: long range base-pairing “A” form tolerates the geometry of G:U base pairs
Can you read ? (the sequence of this DNA)
Sequence-Specific Recognition of DNA by proteins: What do proteins “see” ?
Recognition of Specific sequences by DNA-binding proteins Distribution of H-bonds Donors (D) Acceptors (A) and Hydrophobic groups (H) H Major groove H N O CH3 D N7 A A H A T N1 H N3 N9 Minor groove N1 A A N3 O dR dR Major groove H D O H CH3 N H A A N7 Minor groove T A N1 H N3 N9 A A N1 N3 O dR dR Conclusion: DNA binding proteins can differentiate A-T base pairs from T-A base pairs if they bind from the major groove side, but not from the minor groove side
Patterns of H-bonds Donors (D), Acceptors (A), and Hydrophobic groups (H) available for recognition Recognition of Specific sequences by DNA-binding proteins C H G O H N D N7 Major groove A A N3 H N1 N9 N1 Minor groove N3 A O N H A dR dR D H H Major groove D O H N G A C A N7 N1 H N3 N9 Minor groove A N1 N3 A O H N D dR dR H Conclusion: DNA binding proteins can differentiate G-C base pairs from C-G base pairs if they bind from the major groove side, but not from the minor groove side
DS DNA (Helix) What influences the equilibrium ? (important because DNA is “opened” during replication and transcription) In favor of double-stranded DNA - Hydrogen bonds between strands (minor) - Base stacking Interactions (major) In favor of single-stranded DNA - Electrostatic Repulsion between strands - Entropic considerations: Increased entropy for ssDNAvsdsDNA 2 SS DNAs (“random coils”)
Experimental Studies of DNA denaturation Relative Absorbance Hyperchromic Effect: SS DNA > native DNA DNA molecule Denatured DNA Native DNA 240 180 200 220 260 280 Wavelength (nm) UV spectroscopic analysis of SS (denatured) vs DS (native) DNA “melting curves” for two different DNA molecules (red and blue) show different “melting points” = 2 different Tms
DNA melting is a cooperative process: this explains the sigmoid denaturation curves Increasing Conformational Entropy Increasing Entropy ( 1 -> 2 molecules)
The Tm of a DNA molecule is a linear function of its G-C content
The Tm of a DNA molecule is a linear function of its G-C content/this is not because of higher energy of 3 H-bonds (GC) vs 2 (AT) Effect of G-C content on Stability is due to higher stacking of G-C base pairs compared to AT base pairs DG BP≅ contribution of stacking to the stability of base pair Yakovchuk P et al. Nucl. Acids Res. 2006; 34:564-574