Download

1 / 1
10 likes | 91 Views
Investigating the development of chirality in higher-level organisms, using theoretical models to study protein interactions and RNA involvement in left-right symmetry breaking processes.

E N D
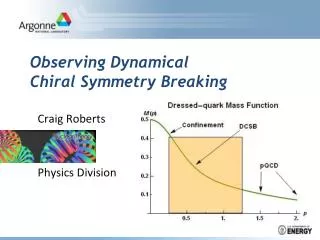
Theoretical Biophysics at UBC,Dr. Steven Plotkin Physics of left-right symmetry breaking Electromagnetism is an achiral force ultimately responsible for the development of an organism, yet essentially all higher level organisms exhibit chirality (most people are right-handed for example). Distance metrics for high-dimensional objects Network of interacting proteins and RNA involved in left-right symmetry breaking. An 8 day old vertebrate embryo (1A ) has a structure more or less where your bellybutton would be (1B) called Henson's node. Increasing the magnification further (6000x) shows that the node is decorated with cilia. These cilia rotate clockwise when viewed from above the node, pushing fluid flow to the left. This is why your heart is on the left (and liver on the right). We study the origins of this symmetry-breaking process. A non-crossing space curve adds complication to the problem, interactions are then non-local, and the path of shortest distance from one configuation to another becomes more complex. “Phase diagram" for the symmetry breaking process, as a function of expression levels. equations governing the protein network. Equation governing the distance between two space curves is a nonlinear PDE with Lagrange functional constraints. A We wish to theoretically model the trajectories of latex beads added to the extraembryonic fluid. Shown here are sample trajectories for healthy (B) and genetically modified (C) mice. This demonstrated that fluid flow was essential for symmetry breaking. Expression levels of RNA and proteins as a function of space and time. Smoothed backbone structure of a protein. we are instested in calculating the "distance" from a random walk configuration to this structure. B C D Steiner problem We are interested in finding the shortest network connecting a set of N points. This is known as the Steiner problem, and it has applications to phylogeny. We are interested in the mathematical properties of Steiner networks in high dimension (~10^6). Protein folding coupled to a thermal ratchet (collaboration w Shirin Hadizedeh & Joerg Rottler ) We study the behavior of a protein which can Spontaneously fold and unfold, in the presence of a thermal ratchet. We are interested in addressing fundamental questions involving apparent violations of the 2nd law of thermodynamics. The nonequilibrium energy in a thermal ratchet can drive a system of particles uphill (A). The drift velocity is a function of the frequency that the ratchet is flashed (B). Thermal ratchets can be used to segregate particles (C). (courtesy Dean Astumian). Steiner Canada. (cf. Berne & Graham Sci Am 1989) (soap-film) A soap-film finds a Steiner network solution to the problem, however the length may not be minimal. Here a local minimum network for some major cities in the US and is found. Steiner network for a collection of 10,000 points randomly distributed in the plane. Protein folding problems We study several problems in protein folding, from dynamics issues, to properties of the Hamiltonian, to structure prediction. Quaternions for modelling DNA-nanopore translocation (Collaboration w Chris Yearwood & Andre Marziali) Rigid bodies modelled by Euler angles show singularities in dynamics exhibited by a divergence in the kinetic energy. This problem is resolved by modelling rigid bodies with hypercomplex numbers (Quaternions). Stereo view (cross your eyes to see a 3-D image) of a partially folded "transition state" structure (blue tube) superimposed on the folded structure of a protein called Chymotrypsin Inhibitor. The increase in the folding nucleation barrier as the Hamiltonian is varied to include more many-body interactions. In order to improve computational efficiency, we coarse-grain the DNA modeled in the simulation using a Gay-Bern-like potential called the RE2 Potential. This potential requires that the bases of the DNA be parameterized as ellipsoids. Euler angles are insufficient for this purpose as they exhibit divergences in their kinetic energy when the ellipsoids are allowed to freely rotate (see figure A: divergences in the Euler angles subsequently cause divergences in the kinetic energy). In order to resolve this issue quaternions, which describe rigid body orientations with four parameters, are implemented and any divergences that would occur using Euler angles in the MD simulation are eliminated (see figure B). Novel algorithms are constructed to find the ground state for a high dimensional system with many local minima. The algorithm shown schematically here stores candidate structures in bins and recalls them later as necessary.