Download

1 / 40
400 likes | 814 Views
Grabbing the Cat by the Tail: Studies of DNA Packaging by Single f 29 Bacteriophage Particles Using Optical Tweezers Acknowledgements Shelley Grimes Dwight Anderson University of Minnesota Sander Tans Douglas Smith Steven B. Smith Yann Chemla Aathi Karunakaran

E N D
Grabbing the Cat by the Tail: Studies of DNA Packaging by Single f29 Bacteriophage Particles Using Optical Tweezers
Acknowledgements Shelley Grimes Dwight Anderson University of Minnesota Sander Tans Douglas Smith Steven B. Smith Yann Chemla Aathi Karunakaran University of California, Berkeley
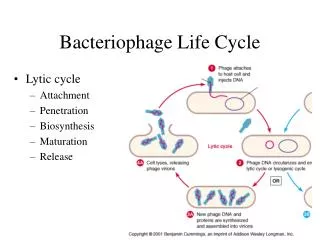
Bacteriophages Icosahedral bacteriophageshave played an central role in the development of Molecular Biology Simplest infectious organisms known Capsid: an empty protein shell that contains the genetic material of the phage Tailand associated protein filaments
Bacteriophage f29 Volume of the capsid: V ~ 56 x 10-3mm3 Length of f29 DNA: 19, 285 bp ~ 6.5 mm
DNA confinement DNA is compacted about 6000x inside the phage head DNA concentration:~ 500 mg/ml Opposing packaging: electrostatic repulsion, bending rigidity, entropy loss, dehydration. DNA must be kept inside the bacteriophage head at significant pressures.
The packaging motor • The head-tail connector (gp 10) Mw = 36 KDa Stoichiometry = dodecamer Structure recently solved
Other views of the connector Top view with DNA model in channel CryoEM reconstruction of Capsid with connector crystal structure fit in Side view: showing two monomers and DNA Ref. Guasch A et al, JMB (2002)
The packaging motor (Cont’d) • The packaging RNA (pRNA) 174 bases (57KDa) , Stoichiometry = 6mer (5mer) (structure unknown)
The packaging motor (Cont’d) • An ATPase (gp 16) Mw = 39 Kda Stoichiometry = most likely 6/phage Structure unknown
The ATPase (gp16) gp16 – DNA dependent ATPase
Optical Tweezers Beam Axis
Objectives Characterize theForce vs. Velocityrelation of a novel motor that may couple rotation to translation. Determine thestall forceof the motor Does aninternal pressurebuild up in the head? If so, how much? Where in the motor cycle does DNA translocation occur? What is the step size of the motor
Constant force feed-back No feed-back Experimental Setup
Packaging at Constant Force Video by Yann Chemla and Aathi Karunakaran
Constant force (5 pN) experiments • Initial packaging rates~ 100 bp/sec. • Pausesare frequent. Ave. pause duration:4 s ± 5 s.Neither the pause duration nor the intervals between pauses are Poisson distributed. Occur more often at higher fillings.
Motor fluctuations Motor Fluctuations Noise Observed rate variations are 5x larger than noise which is ~ 4 bp/s at 1 Hz bandwidth.
Internal Pressure 8 complexes averaged & smoothed A single complex External force = 5 pN • Rate decreases to zero as head fills up • Up to 105 % of the f29 genome is packaged before stalling • An internal pressuremust be building up due to DNA confinement.
Packaging without Force Feedback Video by Yann Chemla and Aathi Karunakaran
Pipette & trap positions fixed Trap & pipette positions fixed ->> LengthForce Motor stalls at high force.
A powerful motor Average stall force =55 pN Max. force meas.> 70 pN
Force-velocity relationship Single complex traces: - Stall force and initial speed vary - Curve shapes are similar Mean traces for 2 fillings: F vs. velocity curves: at1/3 filling at2/3 fillingthe curve is displaced to the left by~ 14pN External Force = 5 pN 1/3 filling 2/3 filling
Force additivity The good overlapobserved by shifting one curve relative to the other suggests that theinternal and external forcesacting on the motoradd. Ext. and Int. forces must be actingat the same pointon motor.
Internal Force No internal force in first half Internal force ~50pN at completion Pressure~6 MPa or 60 kg/cm2
Work done by the motor Work done to package all DNA: 7.5x10-17 J (2x104 kT or 8.2 x 104 pN nm) Available energy per ATP : 120 pN nm Maximum work done per ATP : 37 pN nm (load = 55 pN; suppose step size=2 bp) efficiency ~ 30% (lower bound)
Partitioning the work Total work done by the motor = 8.2 x 104 pN nm (or ~ 20,000 kBTs) Ebending= EIq/2L = kBTP q/2L =2,180 pN nm (~ 530 kBTs) Econfig. loss =900 pN nm (~ 220 kBTs) Therefore, the dominant factor in the work done by the motor appears to be theDNA electrostatic self-repulsion and dehydration
Mechanochemistry of the motor:dependence of rate on [ATP] Force clamp: <F>~7pN 5mM ADP, 5mM Pi
The motor obeys Michaelis-Menten kinetics [T]n (KM)n+[T]n Hill coefficient n=1 V=Vmax 1 ATP hydrolyzed/cycle, no cooperativity between ATPases
F-v relationships for various [ATP] V decreases monotonically vs. F, ATP two regimes F<40pN, F>40pN 5mM ADP, 5mM Pi less force dependence at low ATP
ATP ATP M1 M2 ADP Pi M3 ADP Pi Where is the translocation step? k1 k2 k3 M1 +T <--> M2T --> M3D --> M1 + D k-1 kcat =k2 k3/ (k2 + k3)~ Vmax KM=(k2 +k-1) k3 / k1(k2 + k3) Vmax/KM =(k2 +k-1)/(k1 + k2) At low ATP, v = Vmax[T]/KM, binding is rate limiting: v depends on k1, k-1,k2 , independent of k3 At high ATP, v = Vmax, binding very fast: v depends on k2, k3, independent of k±1 Binding movement step: k1 and k-1 are F dependent Vmax force independent Vmax/KM force dependent KM force dependent 2. Reaction movement step: k2 is F dependent Vmax force dependent Vmax/KM force dependent KM force dependent 3. Release is the movement step: k3 is force dependent Vmax force dependent Vmax/KM force independent KM force dependent binding reaction release (Keller and Bustamante, Biophys. J. 2000)
Force dependence of Vmax, KM Vmax/KM ~ constant KM decreases with force Vmax decreases with force
ATP ATP M1 M2 ADP Pi M3 ADP Pi Translocation coincides with release Our data is consistent with the translocation step coinciding with the release of products of the catalysis Movement step
Step size If noise Dxrms >> step size d, we cannot measure d directly Measure distribution of times spent in a bin of size Dl (which can be >>Dxrms and d) “residence time ” Distribution of residence times is well-defined For an enzyme that performs the steps in a purely random fashion (i.e., its stepping follows Poisson’s statistics) and has one rate-limiting step, this distribution is: 1 (Dl/d-1)! tDl/d-1 tDl/d P(t,Dl/d) = e-t/t, t=d/v Dl, v are known d? <t>=Dl/v, <t2>-<t>2=dDl/v2
Residence times Measure residence time distributions P(t,Dl) vs. [ATP] Fit to distributions to obtain step size d d = 2.15 Extrapolation to [ATP] 0 gives d~2bp
Current questions • What is the organization of the DNA inside the capsid • Does the motor rotate during translocation? • How does the DNA structure affect the activity of the motor? - chargeless DNA - ssDNA • What is the molecular mechanism of energy transduction?
“Once I met a man who grabbed a cat by the tail and learnt 40% more about cats that the man who didn’t” Mark Twain