Download
1 / 19
190 likes | 524 Views
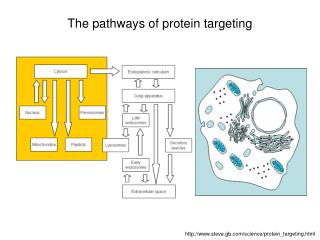
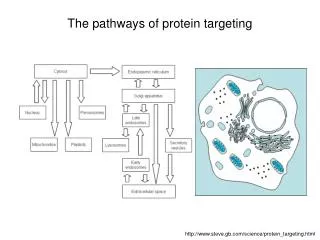
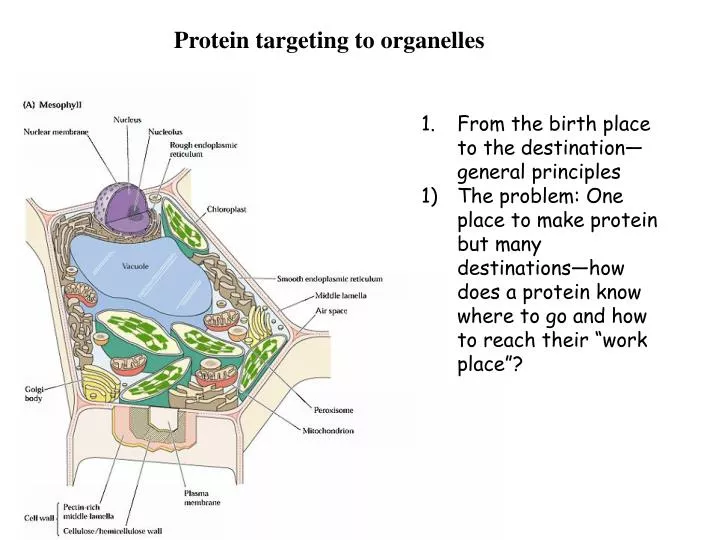
Protein targeting to organelles. From the birth place to the destination—general principles The problem: One place to make protein but many destinations—how does a protein know where to go and how to reach their “work place”?.

E N D
Protein targeting to organelles • From the birth place to the destination—general principles • The problem: One place to make protein but many destinations—how does a protein know where to go and how to reach their “work place”?
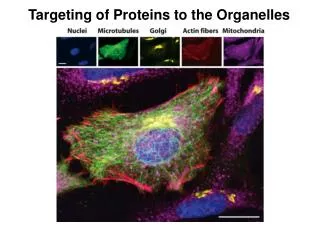
2) How to determine the subcellular localization of a specific protein? The first thing for a research to start is to find out where each protein is working and then find out how they get there. Cell fractionation: traditional Sectioning and TEM immune gold detection Fusion protein: transgenic plants and transient assay
3) Terminology and processes: From cytosol to ER---co-translational transport From cytosol to organelles—post-translational transport (proteins made in the cytosol nucleus, chloroplast, mitochondria, peroxisome, etc) From ER to Golgi to vacuole or plasma membrane and secreted---vesicular transport Signal and receptor: Each targeting process needs a “address label”—peptide sequence on the protein to tell where this protein works. A receptor/machinery needs to recognize that “label/signal” and deliver the protein to the right place. Each protein to be targeted needs to cross at least one membrane barrier (except those remaining in the cytosol).
2. From cytosol to the ER • The first big decision---when a ribosome decides to remain in the cytosol or becomes attached to ER. This determines where a protein will go to work later on. • 2) The signal peptide theory—what makes the first decision • A historical aspect and accidental discovery: from in vitro translation experiment to protein processing, to the role of ER membranes, to the signal peptide as targeting signal for ER attachment. • In vitro labeling of new proteins: Met-S-35 radioactive isotope---run the gel • In the presence of ER: the secreted viral protein was made at right size; but larger when ER is absent • Protease sensitivity assay • Conclusion: secreted protein was translocated into the ER and the size reduction is due to the cleavage of part of the protein that may serve as signal peptide for targeting.
3) The signal and receptor machine: • The signal peptide is often a hydrophobic stretch of 20-40 AA at the N-terminus of the protein. Cleaved after translocation into ER. • The SRP is a protein-RNA complex (1 RNA of 300 base long and 5 proteins). SRP54 is the major component that recognizes and binds the signal peptide. It is a GTPase. Two other SRP proteins work to stop the translation. SRP is normally located in the cytosol but can associate to the membrane by interacting with its receptor in the ER membrane. It was discovered as a peripheral membrane protein by salt wash and reconstitution procedures…
c) The SRP receptor: is an integral membrane protein. Complex of two subunits that are both GTPases. The complex interacts with SRP—as docking site. One of the SRP receptor proteins also interacts with signal peptide. d) The translocon: the protein channel. As the SRP and its receptor interacts, a third protein complex comes into play. This translocon also serves as “receptor” for ribosome that in turn serves as a gating machine to block the channel from leakage of ER content to the cytosol.
4) The process: • The SRP: the “shuttle bus” • SRP receptor: the docking site for the shuttle • Ribosome receptor and the translocon—protein channel • peptidase
5) The topology of membrane proteins in the ER After the signal peptide is removed by peptidase, The soluble proteins are released to the lumen and folded there by chaperones. The transmembrane proteins will be inserted into the membrane during the translocation according to the topogenic sequences inside the protein. After the signal prptide is gone, the first transmembrane domain will serve as “anchor” to stop first translocation of the protein. For multi-transmembrane protein, the first transmembrane domain serves as “stop transfer” and next one serve as “start transfer” signal and this pattern is repeated. Transcolon “moves” to deliver Protein domains across the membrane. ER proteins are born
6) Quality control of ER proteins a) What changes (“education”) are occurring to ER proteins? Similar processes as discussed earlier for posttranslational modifications: Cleavage (of signal peptide), proper S-S formation, glycosylation, folding and assembly b) Quality control: unfolded proteins will be recognized by carrier proteins and transported back into the cytosol for degradation by ubiquitin-dependent proteasome. c) Response to the misfolding of the ER proteins: more chaperone proteins will be made (like in the case of heat shock response).
3. From ER to Golgi • and beyond • --vesicular transport: cargo proteins are transported in a sealed container that is made of membrane vesicle • The road map:
2) From ER to cis-Golgi: • The Golgi complex: three separatedcompartments: • (Close to ER) cis/medial/trans (farther from ER) • b) Which proteins move forward from ER—cis Golgi? Forward traffic • c) How does ER residents find ER destination? The retrograde traffic and ER retention signal KDEL at the C-terminus of the ER lumen proteins (see figure in previous page). KDEL KDEL retrograde traffic Cis-Golgi KDEL KDEL Forward traffic ER
3) From cis to medial and trans Golgi network (TGN): Resident proteins stayed behind and other proteins move forward to the TGN. An important modification in Golgi is O-linked glycosylation following N-linked sugar addition in ER. Different compartments of Golgi (cis/medial/trans) have different enzymes that do different things to the protein passengers. cis medial
4. From TGN to vacuole or Plasma membrane/cell wall • From TGN to lysosome/vacuole: a receptor-mediated pathway: in animals and yeast: the receptor recognize a specific sugar label called mannose-6-phosphate added to the protein in cis-Golgi. The enzyme recognize the specific protein substrates and phosphorylate the mannose-OH group. Such M-6-P serves as ligand for the receptors in the vesicular membrane. M-6-P retrograde traffic vacuole M-6-P M-6-P TGN
2) From TGN to PM/CW: a default pathway All passengers without the M-6-P tag will be deposited to the vesicles that will fuse with plasma membrane and soluble proteins secreted out of the cell. TGN
Is there a retrograde traffic from PM to vacuole? --Sure. Endocytosis in animal cells but in plant cells things look less clear. Endocytosis involves receptors for the cargo. TGN PM