Download
1 / 27
580 likes | 1.7k Views
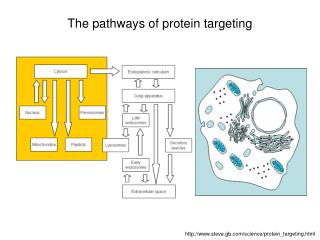
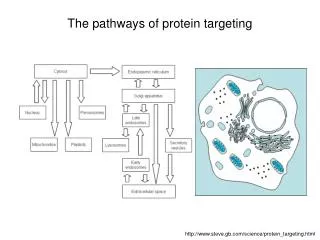
The pathways of protein targeting. http://www.steve.gb.com/science/protein_targeting.html. Protein targeting.

E N D
The pathways of protein targeting http://www.steve.gb.com/science/protein_targeting.html
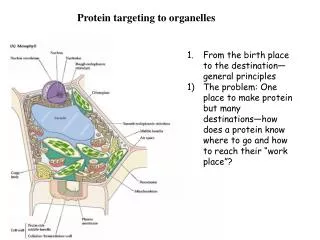
Protein targeting The targeting of proteins is encoded within their structures: signal peptides are stretches (primary structure) of amino acids that target the protein/polypeptide to a particular place, because they are recognised by other proteins whose job it is to import or export proteins into particular organelles. Some signals (so-called signal patches) are formed by tertiary folding, and some signals are effected as oligosaccharides attached to the protein rather than the amino acids themselves.
Translation: Birth of a protein Recall your biochemistry course ... ... or have a look on the 1001 web pages dealing with this topic, .......e.g. http://web.virginia.edu/Heidi/chapter33/chp33.htm
Secretory pathway: Translocation of proteins into the rough endoplasmic reticulum (I) • The synthesis of all proteins begins in the cytoplasm. Proteins destined for the RER (and onwards) have a signal peptide in the first few translated codons, which is recognised by a signal recognition particle (SRP) that pauses translation until the ribosome has been attached to the RER membrane. The SRP contains six polypeptides coating an RNA chain. Prokaryotes have a very similar system to this for the export of proteins out of the cell membrane. • The SRP binds to the SRP receptor (docking protein) on the RER membrane, and directs the nascent polypeptide into a protein translocator (Sec61), which is unplugged as the ribosome is recruited. SRP SRP receptor signal peptide Sec61
Secretory pathway: Translocation of proteins into the rough endoplasmic reticulum (II) • SRP then dissociates and the polypeptide continues to be synthesised, and is threaded through the translocator. Although GTP is required for correct docking, it is not required for translocation. • Finally, a signal peptidase cleaves off the signal peptide, freeing the protein into the ER. signal peptidase
Secretory pathway: Translocation of proteins into the rough endoplasmic reticulum (III) • 'Stop transfer' sequences can be used to thread loops of protein through the RER. • Stop transfer sequences can produce membrane-spanning proteins. • Ion channels often span the membrane 7 or more times. This is achieved by having several start/stop transfer signals. These signal peptides are not consensus sequences, but instead rely on a positive charge followed by hydrophobic stretch followed by polar/negative stretch. The distinction between start and stop transfer signals is largely determined by their order, not by their structure.
Secretory pathway: Translocation of proteins into the rough endoplasmic reticulum (IV)Membrane components http://edoc.hu-berlin.de/dissertationen/unger-christian-2000-03-29/HTML/unger-ch1.html
Secretion: Translocation to the Golgi • After initial glycosylation and quality-control in the RER lumen, most proteins pass into the Golgi. The Golgi has stacked cisternae, which, unlike the RER, are not contiguous. Each cisterna plays a specific role: • * cis Golgi network (CGN) • * cis cisterna • * medial cisterna • * trans cisterna • * trans Golgi network (TGN)
ER to Golgi vesicle flux • (1) Nucleus. • (2) Nuclear pore. • (3) Rough ER. • (4) Smooth ER. • (5) Ribosomes. • (6) Proteins to be transported. • (7) Transport vesicles. • (8) Golgi Apparatus. • (9) cis-Golgi. • (10) trans-Golgi. • (11) Cisternae of the Golgi.
Components of ER to Golgi transport http://www.colorado.edu/MCDB/MCDB1150/ohd/overhead.html The vesicle coat serves to sculpt the curvature of a donor membrane, and to select specific proteins as cargo. It selects cargo proteins by binding to sorting signals. There are three types of vesicle coats: clathrin, COPI (coat protein) and COPII. Clathrin coats are found on vesicles trafficking between the Golgi and plasma membrane, the Golgi and endosomes, and the plasma membrane and endosomes. COPI coated vesicles are responsible for retrograde transport from the Golgi to the ER, while COPII coated vesicles are responsible for anterograde transport from the ER to the Golgi.
Traffic between ER and Golgi • Although most proteins traffic to the Golgi, some are to be retained in the RER. These proteins contain a 'ER return' signal peptide (KDEL) at their carboxy terminus. • H3N+---…---KDEL---COO− • All proteins can traffic into the CGN. However, KDEL receptors (which traffic back and forth) bring escapee proteins back to the ER. The binding of KDEL to receptors is probably pH dependent.
SNARE proteins • SNARE proteins (an acronym derived from "soluble NSF attachment receptor") are a large protein superfamily consisting of more than 60 members in yeast and mammalian cells. • The primary role of SNARE proteins is to mediate fusion of cellular transport vesicles with the cell membrane at the porosome or with a target compartment (such as a lysosome). • SNAREs can be divided into two categories: vesicle or v-SNAREs , which are incorporated into the membranes of transport vesicles during budding, and target or t-SNAREs, which are located in the membranes of target compartments. • Recent classification however takes account of the structural features of the SNARE proteins and divides them into R-SNAREs and Q-SNAREs. • R-SNAREs are proteins that contribute an arginine (R) residue in the formation of the zero ionic layer in the assembled core SNARE complex. Q-SNAREs are proteins that contribute a glutamine (Q) residue in the formation of the zero ionic layer in the assembled core SNARE complex.
SNARE complex formation • Molecular machinery driving exocytosis in neuromediator release. During membrane fusion, the SNARE proteins involved combine to form a SNARE complex. The core SNARE complex is formed by four α-helices contributed by synaptobrevin, syntaxin and SNAP-25, synaptotagmin serves as a calcium sensor and regulates intimately the SNARE zipping.
Secretion: Glycosylation in ER and Golgi • Many proteins are glycosylated in the ER and Golgi. This may either be O-linked (via serine or threonine) or N-linked (usually via asparagine. O-linked glycosylation is characteristic only of proteoglycans, but N-linked glycosylation is performed on all proteins on every asparagine (N) of every NX{ST} sequence in the protein. The core oligosaccharide is composed of two GlcNAc (N-acetylglucosamine) and three mannose residues, but the decoration on this core is very variable. • Glycosylation makes proteins more hydrophilic; it allows them to bind to ECM receptors (lectins); it allows recognition of misfolded proteins by chaperonins; and it causes reduced access to proteases (sterically: oligosaccharides are quite inflexible and restrict access to other molecules). • Glycosylation occurs at the same time astranslocation (indeed, as translation). On the cytosolic face, a precursor oligosaccharide is built from UDP-GlcNAc and GDP-mannose on a membrane-bound molecule of dolichol-phosphate. Dolichol is huge terpenoid lipid, which could (and does) span the RER membrane three times. The dolichol phosphate is then flipped to the lumenal side. On the lumenal side, two oligosaccharide units are preconstructed on another molecule of dolichol from GDP-mannose and GDP-glucose, then added to the main dolichol molecule. The oligosaccharide is finally transferred wholesale from the dolichol phosphate onto the protein by an oligosaccharyl transferase. http://www.steve.gb.com/science/protein_targeting.html
Traffic to lysosomes • The Golgi is not the final destination for most proteins that pass into it. Many proteins are either destined for the ECM (extracellular matrix) or for the endolysosome system. • Lysosomes are the principal site of cellular degradation. They contain hydrolytic enzymes, but are otherwise extremely variable in shape, role and contents. Lysosomes mainly destroy broken organelles and digest endocytosed particles and molecules. The inside of a lysosome is maintained at pH 5 by a proton ATPase and is full of hydrolytic enzymes; consequently these enzymes must be carefully targeted.
Mannose-6-P is a marker for protein targeting to lysosomes • Signal patches mark enzymes destined for the lysosome with GlcNAc-P in the cis Golgi cisterna. The GlcNAc is cleaved off, leaving phosphate attached to mannose of an oligosaccharide. This mannose-6-phosphate is the signal for lysosomal trafficking. GlcNAc-P: N-acetyl glucosamine phosphate
Mannose-6-P is a marker for protein targeting to lysosomes • Mannose-6-P directs vesicles to fuse with late endosomes. A clathrin coat concentrates the mannose-6-phosphate receptor.
Clathrin • COPI and COPII form vesicles without clathrin. Clathrin is required where the trafficking requires physical 'clout. • Adaptins allow clathrin to interact with and concentrate mannose-6-phosphate receptors (and through the receptors, their ligands, i.e. lysosomally targeted proteins). The low pH in endosomes causes the proteins to dissociate from the receptors, which are carried back to the TGN by vesicles. • Clathrin is a hexameric protein that forms cages. It is composed of three heavy chains and three light chains, arranged in a triskelion. The N-terminal knobs protrude into the 'cage' formed when several clathrin molecules link together. Clathrin forms triskelia, which aggregate to form basketworks. • Clathrin forms a basketwork around small vesicles (and is found in patches on larger vesicles). Four (or more) adaptins allow different vesicles to concentrate different receptors (and their ligands). Adaptins recognise FRXY sequences or phosphorylated amino acids in the cytoplasmic tail of transmembrane proteins. Adaptins therefore bind receptors pointing into the extracellular/lumenal compartment to the clathrin which is found in the cytoplasmic compartment. Dynamin is a GTPase which pinches off budding vesicles. Vesicles rapidly lose their clathrin coat after budding: the uncoating of vesicles also requires energy, in the form of ATP (mediated by hsp70 proteins) and calcium.
Golgi to plasma membrane transport COPII coats vesicles trafficking from ER to Golgi. COPI coats vesicles trafficking between Golgi compartments, from Golgi back to ER, from Golgi to cell surface (constitutive secretion), and between endosomes and lysosomes. Clathrin is used when physical clout is required: in concentrating enzymes to traffic to the lysosome, in creating endocytotic vesicles, and in removing membrane from Golgi vesicles as they mature to form secretory vesicles.
Membrane contact and fusion • Vesicles are targeted to specific compartments by SNAREs and Rab. Rab proteins (which are GTPases) dock the vesicles and confer specificity. • SNARE proteins exist in matched pairs and catalyse membrane fusion. • vSNARE - vesicular • tSNARE - target Rab is a membrane-tethered GTPase. It allows specific docking of vesicles to target membranes, via Rab effector proteins. vSNAREs and tSNAREs conspire to fuse the docked vesicle with the target membrane (the tSNARE may require activation by Rab to do this). On fusion of the vesicle, Rab hydrolyses its GTP, and no longer binds the membrane (a hydrophobic 'tail' no longer pokes out of the molecule enough to anchor it to the membrane). Rab-GDP is ferried back to the donor membrane by GDI (GDP dissociation inhibitor), which delivers it to a GEF (guanosine exchange factor), which swaps its GDP for GTP, completing the cycle.
Constitutive and regulated secretion • Constitutive secretory vesicles carry proteins that are continuously secreted by the cell. These vesicles are pinched off from the trans face of the Golgi complex, travel along microtubule tracks, and immediately fuse with the plasma membrane; thus, releasing their contents. Many unrelated proteins can be packaged into a single constitutive secre-tory vesicle. • Regulated secretory vesicles are also derived from the trans face of the Golgi complex; however, they do not fuse with the plasma membrane until the cell is signaled to secrete. These secretory vesicles often contain protein hormones, digestive enzymes and neuropeptides, depending on the cell type. These secretory vesicles are stored in the cytoplasm, where they aggregate and concentrate their contents. For some of these packaged proteins, proteolytic cleavage triggers their activation. Fusion of a secretory vesicle with the plasma membrane, and subsequent secretion of its stored proteins, occurs in response to a signal often associated with an increase in intracellular calcium. The peptide hormone insulin, which is secreted by cells in the pancreas in response to high levels of glucose in the bloodstream, is an example of regulated secretion. The secretion of insulin promotes the uptake of glucose from the bloodstream and the synthesis of glycogen. http://courses.bio.psu.edu/fall2005/biol230weve/tutorials/tutorial6.htm
Endocytosis • The absorption of material (endocytosos) from the outside environment of the cell is commonly divided into two processes: phagocytosis and pinocytosis. • Phagocytosis (literally, cell-eating) is the process by which cells ingest large objects, such as cells which have undergone apoptosis, bacteria, or viruses. The membrane folds around the object, and the object is sealed off into a large vacuole known as a phagosome. • Pinocytosis (literally, cell-drinking). This process is concerned with the uptake of solutes and single molecules such as proteins. • Receptor-mediated endocytosis is a more specific active event where the cytoplasm membrane folds inward to form coated pits. These inward budding vesicles bud to form cytoplasmic vesicles. http://commons.wikimedia.org/wiki/Image:Endocytosis_types.svg
Receptor-mediated endocytosis of LDL • Low density lipoprotein (LDL) is a 500 kDa protein which binds phospholipids and cholesterol. • It binds to LDL receptors localised in clathrin coated pits. LDL is endocytosed by the budding off of the pit, and the vesicle fuses with an early endosome. The LDL receptors return to cell surface by exocytotic vesicular traffic. The early endosome matures, and eventually fuses with or becomes a lysosome. Mutant LDL receptors that cannot bind clathrin cause atherosclerosis.
Endocytotic pathway • Most endocytosed vesicles end up in early endosomes. The low pH (6) inside these organelles releases their ligands. The membrane and receptors are generally recycled back onto the plasma membrane, and the ligand is digested. http://mol-biol4masters.org/Co_and_Post_Translational_Events5-Cellular_Protein_Traffic.htm
Summary: Secretion and Endocytosis http://www.steve.gb.com/science/protein_targeting.html