Download

1 / 58
680 likes | 1.74k Views
Chapter 15 Principles of metabolic regulation: Glucose and glycogen. 15.1 The metabolism of glycogen in animals 15.2 Regulation of metabolic pathways 15.3 Coordinated regulation of glycolysis and gluconeogenesis 15.4 Coordinated regulation of glycogen synthesis and breakdown

E N D
Chapter 15 Principles of metabolic regulation: Glucose and glycogen 15.1 The metabolism of glycogen in animals 15.2 Regulation of metabolic pathways 15.3 Coordinated regulation of glycolysis and gluconeogenesis 15.4 Coordinated regulation of glycogen synthesis and breakdown 15.5 Analysis of metabolic control
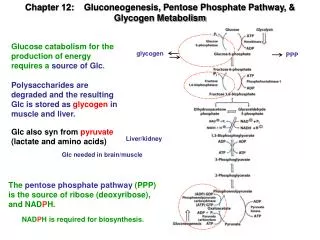
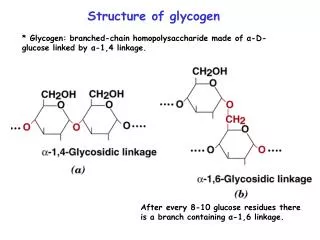
Glycogen synthesis A polymer of glucose Found in liver and muscle Liver: maintain blood glucose conc. Muscle: glycolytic pathway Why so many branches?
Glycogen biosynthesis • Base: Glycogenin with glucose in acetal linkage to Tyr-OH • Glycogen synthase: a(1 4) linkage of glucose from UDP-glucose • UDP-glucose pyrophosphorylase: UTP + G-1-P to UDP-glucose (+PP 2Pi) • Branching enzyme: amylo(1,4 1,6) transglycosylase • Chain moved is 6-7 residues long and from 11 residue chain • Branching point: a(1 6) linkage every 8-12 residues
Fig. 15.11 Fig. 15.7
Glycogen synthesis Fig. 15-18
Branch synthesis in glycogen Fig. 15-9. A segment of 6-7 residues is removed from a branch at least 11 residues long and each new branch point must be at least 4 residues (usually 8-12) away from the nearest existing branch point
SUMMARY 15.1 The Metabolism of Glycogen in Animals • Glycogen is stored in muscle and liver as large particles. Contained within the particles are the enzymes that metabolize glycogen, as well as regulatory enzymes. 2. Glycogen phosphorylase catalyzes phosphorolytic cleavage at the nonreducing ends of glycogen chains, producing glucose 1-phosphate. The debranching enzyme transfers branches onto main chains and releases the residue at the (a1→6) branch as free glucose.
SUMMARY 15.1 The Metabolism of Glycogen in Animals 3. Phosphoglucomutase interconverts glucose 1-phosphate and glucose 6-phosphate. Glucose 6-phosphate can enter glycolysis or, in liver, can be converted to free glucose by glucose 6-phosphatase in the endoplasmic reticulum, then released to replenish blood glucose.
SUMMARY 15.1 The Metabolism of Glycogen in Animals 4. The sugar nucleotide UDP-glucose donates glucose residues to the nonreducing end of glycogen in the reaction catalyzed by glycogen synthase. A separate branching enzyme produces the a(1→6) linkages at branch points.
SUMMARY 15.1 The Metabolism of Glycogen in Animals 5. New glycogen particles begin with the autocatalytic formation of a glycosidic bond between the glucose of UDP-glucose and a Tyr residue in the protein glycogenin, followed by addition of several glucose residues to form a primer that can be acted upon by glycogen synthase.
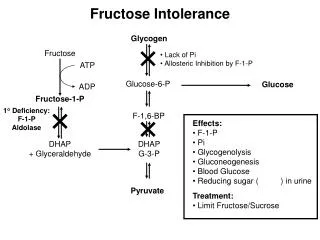
15.3 Coordinated regulation of glycolysis and glucogenesis • Futile cycle ATP + F6P ADP + F1,6BP F1,6BP + H2O F6P + Pi The sum ATP + H2O ADP + Pi + heat
Controls over Enzyme Activity • Six points:(Fig. 15-13) • Rate slows as product accumulates • Rate depends on substrate availability • Genetic controls - induction and repression • Enzymes can be modified covalently • Allosteric effectors may be important • Zymogens, isozymes and modulator proteins may play a role
Fructose 2,6-bisphosphate is a potent regulator of glycolysis and gluconeogenesis • Hormonal regulation of glycolysis and gluconeogenesis is mediated by fructose 2,6-bisphosphate, an allosteric effector for the enzymes PFK-1 and FBPase-1.
Fructose 2,6-bisphosphate in regulation of glycolysis and gluconeogenesis Figure 15.22 Role of fructose 2,6-bisphosphate in regulation of glycolysis and gluconeogenesis. Fructose 2,6-bisphosphate (F26BP) has opposite effects on the enzymatic activities of phosphofructokinase-1 (PFK-1, a glycolytic enzyme) and fructose 1,6-bisphosphatase (FBPase-1, a gluconeogenic enzyme). (a) PFK-1 activity in the absence of F26BP (blue curve) is half-maximal when the concentration of fructose 6-phosphate is 2 mM (that is, K0.5 2 mM). When 0.13 μM F26BP is present (red curve), the K0.5 for fructose 6-phosphate is only 0.08 mM. Thus F26BP activates PFK-1 by increasing its apparent affinity for fructose 6-phosphate. (b) FBPase-1 activity is inhibited by as little as 1 M F26BP and is strongly inhibited by 25 M. In the absence of this inhibitor (blue curve) the K0.5 for fructose 1,6-bisphosphate is 5 M, but in the presence of 25 M F26BP (red curve) the K0.5 is 70M. Fructose 2,6-bisphosphate also makes FBPase-1 more sensitive to inhibition by another allosteric regulator, AMP.
FIGURE 15–23 Regulation of fructose 2,6-bisphosphate level. (Left) The cellular concentration of the regulator fructose 2,6-bisphosphate (F26BP) is determined by the rates of its synthesis by phosphofructokinase- 2 (PFK-2) and breakdown by fructose 2,6-bisphosphatase (FBPase-2). (right) Both enzymes are part of the same polypeptide chain, and both are regulated, in a reciprocal fashion, by insulin and glucagon. Here and elsewhere, arrows are used to indicate increasing (↑) and decreasing (↓) levels of metabolites.
15.4 Coordinated regulation of glycogen synthesis and breakdown • Glycogen phosphorylase is regulated allosterically and hormonally • Glycogen synthase is also regulated by phosphorylation and dephosphorylation • Glycogen synthase kinase 3 (GSK3) mediates the action of insulin (Fig. 15-27) • Allosteric and hormonal signals coordinate carbohydrate metabolism
Fig. 15-24Regulation of muscle glycogen phosphorylase by covalent modification. In the more active form of the enzyme, phosphorylase a, Ser14 residues, one on each subunit, are phosphorylated. Phosphorylase a is converted to the less active form, phosphorylase b, by enzymatic loss of these phosphoryl groups, catalyzed by phosphorylase a phosphatase (PP1). Phosphorylase b can be reconverted (reactivated) to phosphorylase a by the action of phosphorylase b kinase.
Fig. 15-25 Enzyme cascade (Signal transduction cascade) Ca2+ and AMP are allosteric control factors
Fig. 15-26. Glycogen phosphorylase of liver as a glucose sensor When blood glucose returns to normal, glucose binds to phosphorylase a as seen in the figure. When blood glucose is low, glucagon activates phosphorylase b kinase which in turn converts phosphorylase b to its active a form, initiating the release of glucose into the blood.
Chapter 16 The Citric Acid Cycle 16.1 Production of acetyl-CoA 16.2 Reactions of the citric acid cycle 16.3 Regulation of the citric acid cycle 16.4 The glyoxylate cycle
Bridging step ﹛ Occur in mitochondrial
Introduction • Tricarboxylic acid (TCA) cycle aka Citric Acid Cycle, Krebs Cycle • Pyruvate (actually acetate) from glycolysis is degraded to CO2 • Some ATP is produced • More NADH is made • NADH goes on to make more ATP in electron transport and oxidative phosphorylation
檸檬酸 草乙酸 蘋果酸 異檸檬酸 反丁烯二酸 琥珀酸 酮戊二酸
A brief summary of the TCA cycle • Transfer 2 carbon acetyl Co A to four carbon oxaloacetate, dehydration-rehydration to yield isocitrate, two decarboxylation to produce a-ketoglytarate then succinyl CoA……regenerate oxaloacetate. • Carbon enters as acetyl-CoA and exits as CO2 • Produce ATP, NADH, and enzyme-bound FADH2------Oxidative phosphorylation
16.1 Production of acetyl CoAThe bridging step:oxidative decarboxylation of pyruvate to produce acetate • In eukaryotic cells, glycolysis occurs in the cytosol and TCA in the mitochondria
Pyruvate dehydrogenase complex (PDC) • 3 enzymes: pyruvate dehydrogenase, dihydrolipoyl transacetylase, and dihydrolipoyl dehydrogenase. • 5 coenzymes: Thiamine pyrophosphate (TPP), coenzyme A, lipoic acid, NAD+, and FAD • substrate channeling: The intermediates of the multistep sequence never leave the complex, and the local concentration of the substrate of E2 is kept very high. Channeling also prevents theft of the activated acetyl group by other enzymes that use this group as substrate.
Fig. 16.6 As2O3
16.2 Reactions of the citric acid cycleReaction 1: The citrate synthase reactionEntry into the cycle • Acetyl CoA reacts with oxaloacetate in a perkin condensation. • Regulation of CS: • NADH: allosteric inhibitor • Succinyl CoA: feedback inhibition (fifth step product)
Reaction 2: Isomerization of citrate by aconitase • Citrate Isocitrate • Aconitase uses an Iron-Sulfur cluster (Fig. 16-10)
Reaction 3: Isocitrate dehydrogenase -The first oxidation in the cycle • Isocitrate is oxidatively decarboxylated to yield a-ketoglutarate • oxidation to form a-ketoglutarate and NADH • b-decarboxylation to release CO2 and to form a- ketoglutarate
Reaction 4: a-ketoglutarate dehydrogenase complex • A second oxidative decarboxylation to yield succinyl CoA, NADH, andCO2 • 3 enzymes and 5 coenzymes
a-ketoglutarate dehydrogenase complex A second oxidative decarboxylation • This enzyme is nearly identical to pyruvate dehydrogenase - structurally and mechanistically • Five coenzymes used - TPP, CoASH, Lipoic acid, NAD+, FAD • You know the mechanism if you remember pyruvate dehydrogenase
Reaction 5: Succinyl CoA synthetase—A substrate-level phosphorylation • To catalyze succinyl CoA to succinate • A substrate-level phosphorylation • GTP+ADP GDP+ATP
Reaction 6:Succinate dehydrogenase-- An oxidation involving FAD • Catalyze succinate to fumarate • This enzyme is actually part of the electron transport pathway in the inner mitochondrial membrane • has 3 Fe-S clusters
Reaction 7: Fumarase catalyzes trans-hydration of fumarate • Fumarate is hydrated by fumarase to give malate. • The reaction is uncertain.
Reaction 8: Malate dehydrogenase -- completing the cycle • Malate is oxidized to oxaloacetate by malate dehydrogenase • This reaction is very endergonic (+30kJ/mol). • Produce one more NADH • So far, 3 NADH and 1 FADH2 per pyruvate
A summary of the TCA cycle TCA cycle: • Acetyl-CoA + 3 NAD+ + [FAD] + ADP + Pi + 2 H2O 2CO2 + 3 NADH + 3H+ + [FADH2] + ATP + CoASH Glycolysis and TCA cycle: • Glucose + 2 H2O + 10NAD+ + 2[FAD] + 4ADP + Pi 6CO2 + 10 NADH + 10 H+ + 2[FADH2] + 4ATP • NADH = 2.5 ATP, FADH2 = 1.5 ATP So 10NADH + 2[FADH2]=28 ATP
Why is the TCA cycle so complicated • Anabolism • Catabolism
The TCA cycle provides intermediates for biosynthetic pathways • Biosynthetic process 1. Lipid anabolism • provides acetyl-CoA for fatty acid synthesis • ATP-citrate lyase • malic enzyme 2. Anabolism of amino acids and other metabolites • a-ketoglutarate transaminates to glutamate and other amino acids and purine nucleotides • succinyl CoA to porphyrin • oxaloacetate to aspartate • oxaloacetate to PEP---aromatic A.A., formation of 3-PG., gluconeogenesis
The anaplerotic, or “Filling UP,” reaction • Protein may serve an excellent sources of nutrient energy • Catabolism of amino acids provides: succinate, oxaloacetate, fumarate, a-ketoglutarate. • pyruvate (from glycolysis) • acetyl CoA stimulates pyruvate carboxylase
Biotin • Biotin functions as a mobile carboxyl group carrier • Bound covalently to a lysine • The biotin-lysine conjugate is called biocytin • The biotin ring system is thus tethered to the protein by a long, flexible chain • Whenever you see a carboxylation that requires ATP and CO2 or HCO3-, think biotin!