Download
1 / 53
710 likes | 1.58k Views
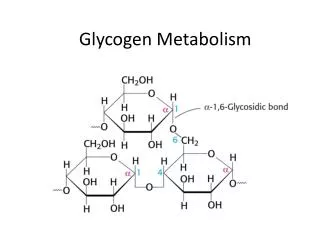
GLYCOGEN METABOLISM. Glycogen Structure. Most of the glucose residues in glycogen are linked by a -1,4-glycosidic bonds . Branches at about every tenth residue are created by a -1,6-glycosidic bonds. Glycogen is an important fuel reserve for several reasons.

E N D
Glycogen Structure • Most of the glucose residues in glycogen are linked by a-1,4-glycosidic bonds. • Branches at about every tenth residue are created by a-1,6-glycosidic bonds.
Glycogen is an important fuel reserve for several reasons • Glycogen serves as a buffer to maintain blood-glucose levels • Especially important because glucose is virtually the only fuel used by the brain. • Is good source of energy for sudden, strenuous activity • Unlike fatty acids, it can provide energy in the absence of oxygen
The major sites of glycogen storage • The liver (10% by weight) • The skeletal muscle (2% by weight) • Glycogen is present in the cytosol in the form of granules ranging in diameter from 10 to 40 nm
Glycogen degradation consists of three steps • The release of G1-P from glycogen • The remodeling of the glycogen substrate to permit further degradation
Glycogen degradation consists of three steps • The conversion of G1-P into G6-P. • It is the initial substrate for glycolysis • it can be processed by the pentose phosphate pathway to yield NADPH and ribose derivatives • it can be converted into free glucose for release into the bloodstream.
Glycogen metabolism is regulated by • Allosterically: • Allosteric responses allow the adjustment of enzyme activity to meet the needs of the cell in which the enzymes are expressed. • Hormones stimulate cascades that lead to reversible phosphorylation of the enzymes, which alters their kinetic properties. • Regulation by hormones allows glycogen metabolism to adjust to the needs of the entire organism.
Glycogen Breakdown Requires the Interplay of Several Enzymes • Four enzyme activities: • one to degrade glycogen, • two to remodel glycogen so that it remains a substrate for degradation • one to convert the product of glycogen breakdown into a form suitable for further metabolism.
GlycogenPhosphorylase:the key enzyme • Cleaves its substrate by the addition of orthophosphate (Pi) to yield G1-P (phosphorolysis) • Catalyzes the sequential removal of glycosyl residues from the nonreducing ends of the glycogen molecule (the ends with a free 4-OH groups)
DG°´ for this reaction is small because a glycosidic bond is replaced by a phosphoryl ester bond that has a nearly equal transfer potential. • Phosphorolysis proceeds far in the direction of glycogen breakdown in vivo because the [Pi]/[G6-P] ratio is usually >100, substantially favoring phosphorolysis. • The phosphorolytic cleavage of glycogen is energetically advantageous because the released sugar is already phosphorylated
Two Remodeling Enzymes • Transferase: • Shifts a block of three glycosyl residues from one outer branch to the other • a-1,6-glucosidase (debranching enzyme) • Hydrolyzes the a-1, 6-glycosidic bond, resulting in the release of a free glucose molecule. • Glucose is phosphorylated by hixokinase (glycolysis)
This paves the way for further cleavage by phosphorylase. • In eukaryotes, the transferase and the a-1,6-glucosidase activities are present in a single polypeptide chain, in a bifunctional enzyme
Phosphoglucomutase • G1-P formed in the phosphorolytic cleavage of glycogen must be converted into G6-P to enter the metabolic mainstream. • This enzyme is also used in galactose metabolism
Serine G1-P G6-P G1,6-BP Serine
Liver Contains G6-Pase, a Hydrolytic Enzyme Absent from Muscle • A major function of the liver is to maintain a near constant level of glucose in the blood. • The liver G6-Pase, cleaves the phosphoryl group to form free glucose and orthophosphate. • This G6-Pase, is located on the lumenal side of the smooth endoplasmic reticulum membrane
GlycogenPhosphorylase Pyridoxal Phosphate integral group of the Enzyme The Pi substrate binding site
In human beings, liver phosphorylase and muscle phosphorylase are approximately 90% identical in amino acid sequence. • The differences result in important shifts in the stability of various forms of the enzyme.
Phosphorylas exists in two states The T state is less active because the catalytic site is partly blocked. The R state, catalytic site is more accessible and a binding site for orthophosphate is well organized.
Phosphorylase Is Regulated by: • Allosteric Interactions: • By several allosteric effectors that signal the energy state of the cell • Reversible Phosphorylation: • responsive to hormones such as: • Insulin • Epinephrine • Glucagon • The glycogen metabolism regulation differs in muscle than in liver because: • The muscle uses glucose to produce energy for itself, whereas the liver maintains glucose homeostasis of the organism as a whole
P A usually active phosphorylase B usually inactive ATP Phosphorylase kinase
Depending on cellular conditions The R and T states of each of the aor b forms are in equilibrium The equilibrium for phosphorylase a, favors the R-state The equilibrium for phosphorylase b, favors the T-state Phosphorylase a differs from b by a phosphoryl group on each subunit
In muscles-Posphorylase b • High AMP, binds to a nucleotide-binding site and stabilizes the conformation of phosphorylase bin the R-state. • ATP acts as a negative allosteric effector by competing with AMP and so favors the T-state. • G6-P also favorsthe T-state of phosphorylase b, an example of feedback inhibition
Under most physiological conditions, phosphorylase b is inactive because of the inhibitory effects of ATP and G6-P. • In contrast, phosphorylase a is fully active, regardless of the levels of AMP, ATP, and G6-P.
Liver Phosphorylase Produces Glucose for Use by Other Tissues • In contrast with the muscle enzyme, liver phosphorylase abut notb exhibits the most responsive T-to-R transition. • The binding of glucose shifts the allosteric equilibrium of the aform from the R to the T state, deactivating the enzyme • Unlike the enzyme in muscle, the liver phosphorylase is insensitive to regulation by AMP because the liver does not undergo the dramatic changes in energy charge seen in a contracting muscle
Phosphorylase kinase in the skeletal muscle: is (abgd)4 • gis catalytic • abd areregultory • dis calmodulin
Muscular activity or its anticipation leads to the release of epinephrine (adrenaline),from the adrenal medulla. Epinephrine markedly stimulates glycogen breakdown in muscle and, to a lesser extent, in the liver. The liver is more responsive to glucagon, a polypeptide hormone that is secreted by the a cells of the pancreas when the blood-sugar level is low. Epinephrine and Glucagon Signal the Need for Glycogen Breakdown
Epinephrine binds to the b-adrenergic receptor in muscle, whereas glucagon binds to the glucagon receptor in liver. • These binding events activate the a subunit of the heteromeric Gs protein. • A specific external signal is transmitted into the cell
Epinephrine also binds to the 7TM a-adrenergic receptor in the liver, which then activates phospholipase C and, hence, initiates the phosphoinositide cascade • The consequent rise in the level of inositol 1,4,5-trisphosphate induces the release of Ca2+ from endoplasmic reticulum stores. • Recall that the dsubunit of phosphorylase kinase is the Ca2+ sensor calmodulin. • Binding of Ca2+ to calmodulin leads to a partial activation of phosphorylase kinase. • Stimulation by both glucagon and epinephrine leads to maximal mobilization of liver glycogen.
Glycogen Is Synthesized and Degraded by Different Pathways • glycogen is synthesized by a pathway that utilizes uridine diphosphate glucose (UDP-glucose) rather than G1-P as the activated glucose donor.
UDP-Glucose Is an Activated Form of Glucose • UDP-glucose, the glucose donor in the biosynthesis of glycogen, is an activated form of glucose. • The C-1 carbon atom of the glucosyl unit of UDP-glucose is activated because its hydroxyl group is esterified to the diphosphate moiety of UDP.
UDP-glucose is synthesized fromG1-P and (UTP) in a reaction catalyzed by UDP-glucose pyrophosphorylase.
This reaction is readily reversible. • Pyrophosphate is rapidly hydrolyzed in vivo to orthophosphate by an inorganic pyrophosphatase. • The essentially irreversible hydrolysis of pyrophosphate drives the synthesis of UDP-glucose.
Glycogen Synthase Catalyzes the Transfer of Glucose from UDP-Glucose to a Growing Chain
glycogen synthase, is the key regulatory enzyme in glycogen synthesis. • Glycogen synthase can add glucosyl residues only if the polysaccharide chain already contains more than four residues. • Thus, glycogen synthesis requires a primer. • This priming function is carried out by glycogenin, a protein composed of two identical 37-kd subunits, each bearing an oligosaccharide of a-1,4-glucose units. • C1 of the first unit of this chain, the reducing end, is covalently attached to the phenolic hydroxyl group of a specific tyrosine in each glycogenin subunit.
How is this chain formed? • Each subunit of glycogenin catalyzes the addition of eight glucose units to its partner in the glycogenin dimer. • UDP-glucose is the donor in this autoglycosylation. • At this point, glycogen synthase takes over to extend the glycogen molecule
A Branching Enzyme Forms a-1,6 Linkages • Branching occurs after a number of glucosyl residues are joined in a-1,4 linkage by glycogen synthase. • A branch is created by the breaking of an a-1,4 link and the formation of an a-1,6 link. • A block of residues, typically 7 in number, is transferred to a more interior site. • The block of 7 or so residues must include the nonreducing terminus and come from a chain at least 11 residues long. • The new branch point must be at least 4 residues away from a preexisting one.
Branching is important because it increases the solubility of glycogen. • Branching creates a large number of terminal residues, the sites of action of glycogen phosphorylase and synthase. • Thus, branching increases the rate of glycogen synthesis and degradation.
Net charge after posphorylation Glycogen Synthase Is the Key Regulatory Enzyme in Glycogen Synthesis • Glycogen synthase is phosphorylated at multiple sites by protein kinase A and several other kinases. • The resulting alteration of the charges in the protein lead to its inactivation • Phosphorylation has opposite effects on the enzymatic activities of glycogen synthase and phosphorylase
Phosphorylation converts the activea form of the synthase into a usually inactiveb form. • The phosphorylatedb form requires a high level of the allosteric activatorG6-P for activity • The a form is active whether or notG6-P is present
Glycogen Is an Efficient Storage Form of Glucose • One ATP is hydrolyzed incorporating glucose 6-phosphate into glycogen -ATP
Glycogen 90% 10% branch G1-P Glucose G1-P -1 ATP -1 ATP G6-P The complete oxidation of glucose 6-phosphate yields about 31 molecules of ATP. Storage consumes slightly more than one molecule of ATP per molecule of glucose 6-phosphate; so the overall efficiency of storage is nearly 97%. +31 ATP Pyrovate
Glycogen Breakdown and Synthesis Are Reciprocally Regulated • By a hormone-triggered cAMP cascade acting through protein kinase A
Protein Phosphatase 1 Reverses the Regulatory Effects of Kinases on Glycogen Metabolism • The hydrolysis of phosphorylated serine and threonine residues in proteins is catalyzed by protein phosphatases. • Phosphatase 1 (PP1), plays key roles in regulating glycogen metabolism.
Insulin Stimulates Glycogen Synthesis by Activating Protein Phosphatase 1 • When blood-glucose levels are high, insulinstimulates the synthesis of glycogen by triggering a pathway that activates protein phosphatase 1
Glycogen Metabolism in the Liver Regulates the Blood-Glucose Level • After a meal rich in carbohydrates, blood-glucose levels rise, leading to an increase in glycogen synthesis in the liver • Insulin is the primary signal for glycogen synthesis • The liver senses the concentration of glucose in the blood, (~80 to 120 mg/100ml). • The liver takes up or releases glucose accordingly.
The amount of liver phosphorylase adecreases rapidly when glucose is infused. • After a lag period, the amount of glycogen synthase aincreases, which results in the synthesis of glycogen.
Phosphorylase a is the glucose sensor in liver cells. • The binding of glucose to phosphorylase ashifts its allosteric equilibrium from the active R form to the inactive T form. • This conformational change renders the phosphoryl group on serine 14, a substrate for PP1. • It is significant that PP1 binds tightly to phosphorylase a but acts catalytically only when glucose induces the transition to the T form. • Recall that the R-T transition of muscle phosphorylase ais unaffected by glucose and is thus unaffected by the rise in blood-glucose levels