Download

1 / 74
740 likes | 1.01k Views
RNAi Overview Mechanism Role in heterochromatin formation Regulation of transcriptional networks Applications. Molecular Biology’s Central Dogma. DNA. AAAA. RNA. mRNA rRNA tRNA. protein. other noncoding RNAs. RNAi :.

E N D
RNAi • Overview • Mechanism • Role in heterochromatin formation • Regulation of transcriptional networks • Applications
Molecular Biology’sCentral Dogma DNA AAAA RNA mRNA rRNA tRNA protein other noncoding RNAs
RNAi: • RNAi is a powerful, conserved biological process through which the small, double-stranded RNAs specifically silence the expression of homologous genes, largely through degradation of their cognate mRNA. • Endogenous cellular mechanisms • Effector molecules for functional genomics • Great potential as therapeutic agents for treatment of human disease
dsRNA intermediate inhibitsprotein translation or causes mRNA degradation (miRNA) cleavage of targeted mRNA (siRNA) RNAi is mediated by small (~21-25 nucleotide) noncoding RNAs complementary to the targeted gene mRNA: AAAA
History of RNA-mediated suppression Curious findings in plants and worms showed the way… 42% Napoli C. et al, Plant Cell, 1990 Post-transcriptional gene silencing in petunias engineered to overexpress pigment gene (chalcone syn) reversible co-suppression of homologous genes in trans
Another strange phenomenon Guo and Kemphues - 1995 Par-1 gene in C. elegans Inject antisense or, as a control, sense RNA to Par-1 to test the function of the gene. Both sense and antisense gave the same phenotype, but non-specific RNAs gave no phenotype??? 1st evidence of dsRNA gene silencing
Small double-stranded RNAs mediate silencing in nematode negative control uninjected Fire and Mello antisense mex-3 dsRNA mex-3 Fire et al (1998). Potent and specific genetic interference by double-stranded RNA in C. elegans. Nature 391:806 DOUBLE STRANDED RNA MORE EFFECTIVE THAN EITHER STRAND INDIVIDUALLY
RNAi works in Drosophila as well as C. elegans
Breakthrough in mammalian gene silencing: engineered small interfering RNAs (siRNAs) Lamin siRNA LacZ siRNA buffer siRNAs corresponding to a targeted gene can suppress its expression with few nonspecific effects. Elbashir et al. (2001) Nature 411:494
Longstanding problem in mammalian cells: nonspecific suppression induced by long dsRNA
Why do organisms have RNAi machinery? Serves, in part, as a cellular defense role against foreign RNAs (innate antiviral defense mechanism in nematodes, insects and plants)
RNAi • Overview • Mechanism • Role in heterochromatin formation • Regulation of transcriptional networks • Applications
Features of RNA interference (RNAi) • dsRNA works significantly better than either sense or antisense RNA alone • dsRNA must be targeted to exon sequences intron and promoter sequences do not produce detectable interference • Introduction of dsRNA produces a pronounced decrease or elimination of the endogenous mRNA transcript • Only a few dsRNA molecules are necessary to elicit a systemic response that is passed on to progeny • dsRNA interference can cross cellular boundaries 2 nt 3’ overhang G/C content <50% Blast searching to exclude complementarity How does it work?
Small RNA biogenesis in animals: siRNA and miRNA Tomari and Zamore (2005) Perspective: Machines for RNAi. Genes Dev 19:517
step one: 34 27 21 20 16 short-interfering RNA processing the dsRNA into 21-23 nt fragments Tuschl, 2001
What cleaves the dsRNA into 21-23 nt fragments? RNase III enzymes were previously known to cleave both strands of dsRNAs to leave 2 nt 3’ overhangs The same types of products formed when the dsRNA is degraded The Drosophila genome appears to encode 3 classes of RNase III enzymes RNase III - RNC_CAEEL Drosha 2 RNase III domains, 1 helicase domain (CG4792 and CG6493)
CG4792 protein can generate siRNAs CG4792 was renamed Dicer-1 CG6493 = Dicer-2 Are RNase III enzymes involved in siRNA synthesis? (cleavage of ribosomal RNA (rRNA) precursors) Transfect Drosophila S2 cells with T7 epitope tagged-RNase III expression vectors Immunoprecipitate with T7 antibodies Test whether immunoprecipitates can generate 21-23 nt fragments from dsRNA
Dicer contains two RNAse III domains long dsRNA siRNAs
19 nt duplex 2 nt 3’ overhangs siRNAs have a defined structure
step two: the antisense strand of the siRNA guides cleavage Tuschl, Cell, 2002
RNAi silencing complex (RISC) • may be associated with translating ribosomes • active RNAse enzyme (Slicer) identified ? • may participate in endogenous pathways
Post-transcriptional gene silencing (PTGS) “siRNA” molecule: “Dicer” processes long dsRNA into short siRNA: “Guide” (antisense) strand incorporated into RISC complex: guides RISC to complementary sequences in target mRNAs (RNA-induced silencing complex) Loaded onto Ago2 Nat Rev Genet 3:737 (2002)
Argonaut protein (AGO2) = endonuclease“Slicer” Originally described in plant development and for germline stem-cell division in flies
Ago2 is the sole “Slicer” of the Ago family, essential for mammalian development Ago2 = RNAi Ago1 = miRNA-med gene silencing Liu et al (2004) Argonaute 2 is the catalytic engine of mammalian RNAi. Science 305:1437
Argonaute structure All eight human AGO family members conserve the PAZ-PIWI domain Paz: 3’ guide strand binding PIWI : RNase H-like active site and 5’ guide strand binding Three groups: Ago-like proteins, Piwi-like (related to Drosophila P-element induced wimpy testis) and new Group 3 Agos.
RISC assembly pathway for siRNA. How is the guide strand selected? Tomari and Zamore (2005) Genes Dev 19:517
Kim et al, Nat Rev, 2007 Mechanisms of RNA interference in mammalian cells Engineered siRNAs enter pathway here Note: miRNAs can have complete seq complementarity = cleavage e.g. miR-196-directed cleavage of Hoxb8 Processing body Global inhibition of miRNA biogenesis decreases #/size of P bodies
Exportin-5 mediates the nuclear export of pre-microRNAs and short hairpin RNAs Rui Yi, Yi Qin, Ian G. Macara and Bryan R. Cullen Genes & Development (2003) 17:3011-3016 Wan-Ting Chen
RNAi • Overview • Mechanism • Role in heterochromatin • Regulation of transcriptional networks • Applications
MicroRNAs: naturally occurring RNAi molecules Novel class of genes that regulate gene expression during development. Play important roles in cellular and developmental processes. miRNAs regulate gene expression by base pairing imperfectly to the 3’-UTR of target mRNAs and inhibiting protein synthesis or causing mRNA degradation. Fine tuning of gene expression ~30% of all genes subject to regulation by miRNAs
let-7 interacts with the 3’ UTRs of several mRNAs lin-4 is another small RNA that interacts with the 3’UTRs of mRNAs Fusing the lin-41 3’UTR to the lacZ ORF causes lacZ expression to be regulated in a let-7 dependent manner
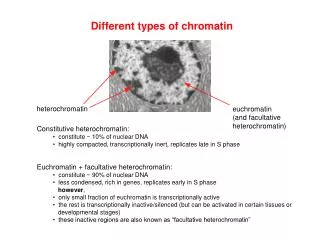
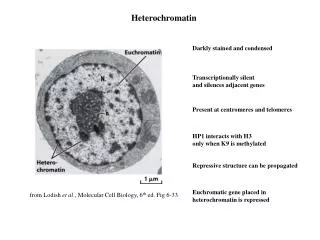
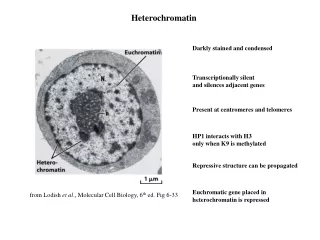
Heterochromatin • 1928, staining differences in nuclei lead to terms euchromatin and heterochromatin • Heterochromatin • Dark staining • Genetically inactive • Few genes, those present repressed • Replicates late in S phase • Centromeres and telomeres are heterochromatic • Portion of Y and inactivated X chromosomes • Position effect when genes are translocated to location adjacent to heterochromatin
Chromosome Banding of Mitotic Chromosomes • C-banding • Giemsa stain only stains centromeric regions • G-banding • Trypsin treatment before Giemsa staining • Provides a unique series of bands along each chromosome • Provided uniform nomenclature for human chromosomes/locations in 1971
Heterochromatin formation: Transcriptional gene silencing (plant/fungi).
Transcriptional Gene Silencing (TGS) RNA-directed DNA Methylation RNA-induced Transcriptional Silencing (RITS) Induction of H3K9 methylation HP1 recruitment Heterochromatin spreading
Heterochromatin and RNAi AreRequired to Establish CENP-AChromatin at Centromeres Hernan Diego Folco, Alison L. Pidoux, Takeshi Urano, Robin C. Allshire Hsiang Ho
otr : outer repeats imr : innermost repeats cnt/cc : central core otr otr Background • Centromere nucleosome differs from conventional nucleosome by the presence of a centromere-specific histone H3 variant (CENP-A, centromere protein A, in place of canonical H3) • CENP-A cnp-1 = fission yeast homologue of CENP-A • In fission yeast, kinetochore establishment requires flanking heterochromatin providing assembling signal. These could include RNA-induced transcriptional silencing (RITS), H3K9Me, HP1, cohesin. • siRNA originating from otr serves as platforms for the recruitment of modifying enzs, direct H3K9Me to initiate heterochromatin formation However, the primary signals specifying the site of CENP-A chromatin, and thus kinetochore assembly, are unknown…
Hypothesis Heterochromatin marks sites for CENP-A chromatin assembly Experimental system Endogenous chromosome are tethered by cohesin, so to study the mitotic stability of centromere, small circular minichromosomes were used. Clr4: H3K9 methyltransferase CENP-A chromatin formation in clr4Δ is dependent on the entering state: preassembled (cross) or naked DNA (transform)
Fig 1. Clr4 H3K9 methyltransferase and otr heterochromatin are required to establish CENP-ACnp1 chromatin
Fig 2. • CENP-CCnp3 and Sim4 (Kinetochore component) were retained in endogenous cc1/3, cc of pH-icc3i-H and when it is crossed into clr4Δ, but not when transformed into clr4Δ. • Small pH-cc2 or pHH-cc2 minichromosomes may be particularly sensitive to the absence of heterochromatin • ∴minichromosome containing two nearly complete otr repeats flanking an entire cen3 central domain were used. • Same conclusion were reached as in pH-cc2 or pHH-cc2 . ∴kinetochore proteins association depends on initial propagation • Once CENP-ACnp1 chromatin and kinetochore proteins assemble on centromeric DNA, they are propagated without flanking heterochromatin. Clr4 is required to establish, but not to maintain CENP-ACnp1 and kinetochore proteins.
Fig 3. clr4+ reintroduction triggers CENP-ACnp1 recruitment to minichromosome (A) and minichromosome stability (B). • Not sensitive to TBZ (white)= fully restore centromere function. Red =lost • Clr4 reintroduce restore full centromere-kinetochore function Clr4 reintroduce bring back CENP-ACnp1 of clr4Δ-T
Fig 4. Swi6 and RNAi components are required for CENP-A establishment. RNAi-mediated otr heterochromatin assembly =HP1 siRNA originating from otr serves as platforms for the recruitment of modifying enzs, direct H3K9Me A patch of fully intact centromeric heterochromatin is required to establish centromere
Key findings • RNAi-directed heterochromatin flanking the central kinetochore domain at fission yeast centromeres is required to promote CENP-A cnp-1 and kinetochore assembly over the central domain. Figure S6. Model. An intact domain of heterochromatin with all components (RNAi, Swi6, Clr4 H3K9me2) is required for the initial establishment of CENP-ACnp1 chromatin and consequently kinetochore proteins on central domain DNA. However, once established, this specialised CENP-ACnp1 structure can be propagated independently of heterochromatin.
Questions raised • Fig 3B, clr4Δ-X is also sensitive to TBZ. This indicates that although CENP-ACnp1 is formed (IP positive), but the centromere function can still be impaired. • Fig 4, in the absence of chp1, dcr1, or swi6, CENP-ACnp1 did not associate with cc2 on minichromosome even though itis associated with endogenous centromere. This indicates either RNAi is required on naked DNA or it might be other components are defected in the artificial minichromosome (ex.cohesin). Crossing to mutants may address the difference is due to the fact that DNA is naked or preassembled.
Linkage of H3K9 methylation and HP1 to regulation of euchromatic genes • Reversible silencing of euchromatic genes (Rb) • Marks found with transcribed chromatin • Ex: rolled gene in Drosophila located deep in heterochromatin, however expression is essential for viability. Location dependent • Pol II can associate with silent chromatin • Some promoters within heterochromatic domains drive transcription, however resulting RNA is probably degraded – important for continued silencing