Download

1 / 54
540 likes | 783 Views
Fundamental Features of Eukaryotic Gene. Eukaryotes VS Prokaryotes. Key differences between eukaryotic and prokaryotic genomes: Eukaryotic genomes are larger. Eukaryotic genomes have more regulatory sequences. Much of eukaryotic DNA is non-coding. Eukaryotes have multiple chromosomes.

E N D
Eukaryotes VS Prokaryotes Key differences between eukaryotic and prokaryotic genomes: • Eukaryotic genomes are larger. • Eukaryotic genomes have more regulatory sequences. • Much of eukaryotic DNA is non-coding. • Eukaryotes have multiple chromosomes. • In eukaryotes, translation and transcription are physically separated which allows many points of regulation before translation begins.
Eukaryotes VS Prokaryotes • Some eukaryotic genes that have no homologs in prokaryotes: • Genes encoding histones • Genes encoding cyclin-dependent kinases that control cell division • Genes encoding proteins involved in processing of mRNA
Eukaryotes VS Prokaryotes • Gene characteristics not found in prokaryotes: • Eukaryote genes contain non-coding internal sequences (introns). • Form gene families—groups of structurally and functionally related genes.
Eukaryotic Genes Eukaryotic genes have a promoter to which RNA polymerase binds and a terminator sequence to signal end of transcription. Terminator sequence comes after the stop codon. Stop codon is transcribed into mRNA and signals the end of translation at the ribosome.
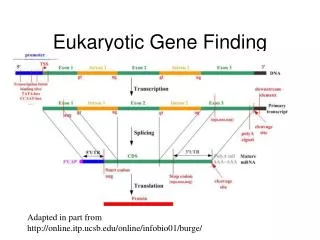
Eukaryotic mRNA Splicing • Researchers had expected the complementary of upstream (5’) and downstream (3’). • However, this idea was soon discounted. • The sequences at 5’ and 3’ are not complementary, yet not random. • The sequences at 5’ ends of introns were similar to each other, as were the sequence at the 3’ ends.
Eukaryotic mRNA Splicing • Almost all introns begin with G:U and end with A:G. • The branch site: sequences within introns, 20-50 nucleotides just upstream of 3’, is essential for splicing. • The consensus sequence of the branch site is YNYURAC. The A is the key nucleotide in the sequence. Mutations of this branch site A prevent splicing.
Splicing is carried out by a complex of proteins and RNA called the spliceosome. • Small nuclear RNAs (snRNAs), U1, U2, U4, U5, and U6 associate with proteins to form small nucleoprotein particles (snRNPs). • The U1 snRNP is the first component to bind 5’ splice site, followed by U2 snRNP binding to the branch site. • The other three snRNPs bring 5’ and 3’ splice site together. The U6 snRNP removes the intron and joins the exons.
Eukaryotic mRNA Splicing Video porpax.bio.miami.edu/.../150/gene/mol_gen.htm
Alternative mRNA Splicing • Many primary gene transcripts can be spliced in different ways to produce distinct RNA molecules that each encode a different protein. • Alternative splicing often produces two forms of the same protein that are necessary at different stage of development or in different cell types. • Immunoglobulins of the IgM class exist as either a membrane bound protein displayed on the cell surface (B cell) or as a soluble protein secreted into the blood (plasma cell).
Alternative mRNA Splicing • Alternative splicing can be extremely complicated, e.g. the gene encoding the protein α-tropomyosin containing 14 exons. • Different combinations of exons are used to form mature tropomyosin mRNAs in different cell types. • The overall structure of each tropomyosin protein is similar, the cell-type specific amino acids may function as binding sites for different proteins.
Alternative mRNA Splicing • The complexity of an organism is not reflected in the number of its genes. • The analysis of human genome identified 22,000 genes, whereas the number of gene transcripts is 35,845 (about the same as Caenorhabditiselegans and less then Arabidopsis thaliana.) • The difference between these two numbers is accounted for by alternative splicing. • One-half of the transcripts are non-coding RNAs.
Errors in Splicing Cause Disease • At least 15% of human genetic disorders are found to result from splicing defects. • Mutation in consensus splicing sequences may lead to exon skipping, deletion of part of exon, and the inclusion of the sequence that should not be part of the mature mRNA. • Mutations in the splicing sites of the β-globin gene disrupt splicing and cause β-thalassemia. • Mutations in (exonic splicing enhancers) ESEs can have profound effects, even a silent mutation can disrupt ESE and cause splicing problem.
Functions of Introns • Introns do not encode proteins, yet maintained in eukaryotic genomes. Why? • It is suggested that introns might facilitated genetic variation (the substrate for natural selection) if exons encode functional domains of proteins and introns promote recombination of exons (exon shuffling.) • The low-density-lipoprotein receptor (LDL-R) appears to be made up of bits of other proteins stitched together to make a new protein.
Function of Introns Evidence of exon shuffling in the LDL-R gene
Functions of Introns • Almost all of the genes for small nucleolar RNA genes (snoRNAs) that are involved in ribosomal RNA maturation in vertebrates are found within the introns of genes that code for proteins. • After splicing and excision of the introns, nucleotides are removed from the 5’ and 3’ ends of introns to produce functional snoRNAs. • The human U22 snoRNA is in an intron of the U22 host gene (UHG) that does not appear to code for a protein. • It is the intronic sequences of UHG that are “useful”.
Genes Are Within Other Genes • Genes can be found within the introns of other genes. • In Drosophila, unrelated gene for a pupal cuticle protein was found embedded within one of the introns of GART gene, coding for enzyme important for biosynthesis of purines. • Intron 22 of the human factor VIII gene is very large (32 kb) and harbors two genes, F8A and F8B.
Genes Are Within Other Genes • The tumor suppressor genes p16INK4a and P19ARF are encoded by a single locus in the human genome. • The reading frames are different resulting in completely unrelated proteins.
Repetitive DNA Sequences • Most genes are present only once per haploid genome. However, the genes for histones, tRNAs and rRNAs are present many times within the genomes and are often clustered together. • Moderately repetitive sequences (genes): code for tRNAs and rRNAs • These molecules are needed in large quantities; the genome has multiple copies of the sequence.
Repetitive DNA Sequences • Four different rRNAs • 16S, 5.8S, 28S are transcribed as a single precursor molecule. Humans have 280 copies of the sequence on five different chromosomes • 5S (S = Svedberg unit)
Repetitive DNA Sequences • The histones gene family consists of major genes (H1, H2A, H2B, H3, and H4). In Drosophila, these 5 genes occur in a cluster of about 5000-6000 bp, and each cluster is tandemly repeated between 100 – 1000 times. • In higher eukaryotes, a cluster of histones genes exists at only 10-40 copies per genome.
Repetitive DNA Sequences • Eukaryote genomes have two types of highly repetitive sequences that do not code for proteins • Minisatellites: 15–100 bp long, repeated 20-50 times. • Microsatellites: 2-6 nucleotides, and are present in tandem arrays of five to about 30 copies. • Number of copies varies among individuals—provides molecular markers.
Repetitive DNA Sequences • Other moderately repetitive sequences can move from place to place in the genome - transposons • Transposons make up 40 percent of human genome, only 3 - 10 percent in other sequenced eukaryotes • Transposons are not tandemly repeated but, rather, exist as isolated elements that may be present in many thousands of copies per genome. • SINEs (short interspersed elements) range in length from 130 - 300 bp; 15 percent of human DNA. One, Alu, is present in a million copies
Gene Duplication and Divergence • Eukaryotic genomes increase in size and complexity through gene duplication and subsequent sequence divergence. • Duplication of a gene allows duplicated copy undergo mutation without selection, because the other copy supplies the protein needed for cell function. This process is called genetic drift. • The evolutionary significant of genetic drift is that mutations may lead to a protein acquiring new functions – an enzyme acting on different substrate or cell type.
Gene Duplication and Divergence • There are 7 genes in the Arabidopsis related to the terpenesynthases, three of which are closely related. • Two genes, 25820 and 25830, are identical and the third gene, 25810, is 80% identical to these two genes. • The 25810 is expressed exclusively in roots and does not synthesize one of the terpenes made by 25820 and 25830. • An ancestral gene underwent duplication; one gene diverged in expression and function to give rise to 25810, whereas the other underwent a second duplication to produce 25820 and 25830.
Gene Duplication and Divergence • Genetic drift in duplicated genes may be desirable since generating genetic diversity. • However, some situations, it needs to be counteracted. • Mutations may be corrected in two ways, by elimination and gene conversion. • For example, unequal crossing-over can lead to the accumulation of extra copies of tandemly repeated genes. These individuals carrying these copies may be selectively disadvantage and will be eliminated from the population.
Gene Duplication and Divergence • Gene duplication and divergence can lead to pseudogene. • DNA sequences are highly related to functional gene, containing inserted or deleted sequences or other mutations that prevent the production of functional protein. • Processed pseudogene - arisen by reverse transcription of an mRNA molecule into DNA.
Gene Regulation in Eukaryotes • Eukaryotes have three different RNA polymerase complexes, making it possible for three independently regulated families of promoters. • The eukaryotic RNA polymerases require more transcription factors to initiate RNA synthesis. • These transcription factors play many different roles including recognizing other transcription factors, removing chromatin proteins that block polymerase access, and unwinding the DNA at the promoter. • Control of eukaryotic gene expression is more complex than in prokaryotes.
Transcriptional Factors • A eukaryotic promoter consists of TATA box, CAT box, and GC box that lie about -25, -75, and -90, respectively. • Each eukaryotic structural gene has its own set of response elements. • In addition to DNA-protein interaction, protein-protein associations are important for regulating eukarytic transcription.
Transcriptional Factors • Formation of an RNA polymerase II transcription initiation complex at a TATA box. • TFIID binds to a TATA box, and in sequence, other transcription factors and RNA polymerase is responsible for initiating transcription.
Transcriptional Factors • Transcription factors must bind DNA and interact with RNA polymerase. • These two functions – DNA binding and activation are carried on different surfaces of the protein (separate domains.) • Domain swap experiment strongly suggested that the DNA binding region might be separate from the transcriptional activating region.
Transcriptional Factors • A transcription factor can activate different targets depending on what other transcription factors are present. • Various combinations of transcription factors are required for transcription.
Eukaryotic Enhancers • The DNA sequence upstream and downstream of even-skipped are termed enhancers, they can enhance transcription of a promoter. • Enhancers contain DNA sequence that are recognized and bound by transcriptional factors. • The transcriptional factors, then, promote transcription by recruiting RNA polymerase and associated factors necessary for transcription.
Eukaryotic Enhancers • Enhancers increase gene expression independently of their position relative to the gene. • Enhancers acts over long distances, as far as 1,000,000 bases away from the promoter. • Enhancers can act at the great distances by being brought in close proximity to a promoter by looping of the DNA strand. • An insulator sequences can block the action of enhancers. • Flanking a transgene with insulators sequences frequently lead to higher levels of transgene transcription.
Eukaryotic Enhancers biology.kenyon.edu/.../Chap10/Chap10.html
DNA Packaging and Gene Expression • DNA molecules are tightly wrapped around histone proteins to form a structure called nucleosomes. • The string of nucleosomes are further looped and wrapped into a compact structure called chromatin. • The “active” chromatin (e.g., that making rRNA) lacked the normal appearance of chromatin. • The actively transcribed DNA, looping out from the chromosome and contain either naked DNA or single nucleosome, is more accessible to nuclease such as restriction enzymes than tightly condensed inactive chromatin.