Download

1 / 50
520 likes | 972 Views
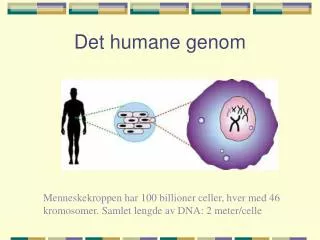
Genom. Alexandr Sember. Genom. Definice: Původní : úplný soubor veškeré genetické informace organismu Novější, přesnější : celková sekvence NK (DNA/RNA), která má informační hodnotu a je charakteristická pro daný organismus. Struktura genomu obecně: Lineární/kružnicová DNA

E N D
Genom Alexandr Sember
Genom Definice: Původní: úplný soubor veškeré genetické informace organismu Novější, přesnější: celková sekvence NK (DNA/RNA), která má informační hodnotu a je charakteristická pro daný organismus Struktura genomu obecně: Lineární/kružnicová DNA Segmentovaná/nesegmentovaná DNA/RNA ss (single-stranded)/ds (double-stranded)
Struktura genomu Struktura genomu konkrétně: Prokaryota: obvykle 1 kružnicová (nebo lineární) molekula (nukleoid) volně v cytoplazmě + plazmidy (kružnicové nebo lineární) Eukaryota: genom segmentovaný na jednotlivé chromozomy, jádro odděleno od cytoplazmy + v cytoplazmě mitochondriální DNA (kružnicová) + někdy chloroplastová DNA (lineární!!) + někdy plazmidy (př. u kvasinek) Archae: cirkulární dsDNA rozdělené u řady zástupců do více molekul, chybí jaderná membrána
Struktura genomu virů Segmentovaný x nesegmentovaný; DNA x RNA; lineární x kružnicový + ssRNA viry (hepatitida A, dětská obrna, klíšťová encenfalitida, SARS, zarděnky…) - ssRNA viry (chřipka, spalničky, příušnice, vzteklina…) dsRNA viry (Reoviridae)…případně viroidy, virusoidy (nejsou viry) Retroviry(ssRNA do dsDNA, reverzní transkripce; př. HIV) + DNA retroviry (př. hepadnaviry – př. hepatitida B) ssDNA viry (parvoviry – onemocnění zvířat, př. psů a koček, někteří bakteriofágové – M13, ΦX170) Virus vztekliny dsDNA viry(papillomaviry – bradavice; většina bakteriofágů; adenoviry – onemocnění dýchacích cest; herpesviry – plané neštovice, pásový opar) Počet genů: 3 – nejmenší RNA viry (bakteriofág MS2) 9-11 – nejmenší DNA viry (ΦX174) cca 150 – největší DNA viry (bakteriofág T2)
Influenza (Orthomyxoviridae) – virus chřipky – minus ssRNA, 8 segmentů, 8 genů, 10 proteinů HIV (Human Immunodeficiency Virus, Lentiviridae) – retrovirus, 2 plus ssRNA, 9 genů (nejen gag, pol, env), 15 proteinů RNA viry rychle mutují!!! – RNA polymeráza nemá korekční aktivitu
Struktura genomu prokaryot Nukleoid svinutý do 30-100 smyček okolo středu; RNA drží smyčky u sebe, proteiny drží nadšroubovicové závity, neutralizace náboje DNA: H1, Hu proteiny, kationty, polyaminy… E. coli: DNA 1,5 mm, průměr buňky 1 um Genomdo 5 Mbp, 2500 – 3500 genů, málo nekódujících sekvencí Plazmidy – obvykle neesenciální geny, rezistence na ATB, pro konjugaci (F plazmid), syntéza toxinů zabíjejících bakterie (Col plazmidy), patogenita (Ti plazmid u Argobacterium tumefaciens, transgenoze). Borrelia burgdorferi(hlavní lineární genom + 17 lineárních nebo cirkulárních plazmidů, nesou i esenciální geny) X Treponema pallidum (příbuzný druh, ne) Vibrio cholerae – 2 cirkulární chromozomy; A. tumefaciens – 3 cirk. + 1 lineární
Struktura genomu eukaryot Genomlineární, segmentovaný na chromozomy Myrmecia pilosula (♀2n=2/♂1n) Nejméně chromozomů Nejvíce chromoz. Ophioglossum reticulatum (2n=96x=1440) Člověk: DNA dlouhá 2 metry (3 x 109 bp x 2 pro diploidní genom x 0,34 nm vzdálenost mezi bp)se musí vejít do buňky velké 10-15 um – nutnost mnohonásobné spiralizace; 2n = 46 chromozomů; záporný náboj DNA odstíněn histony (bazické proteiny); řada dalších komponent chromatinu…
Srovnání prokaryota/eukaryota • Prokaryota: • Malé, kompaktní genomy, téměř samé geny • Vyjímečně mají introny v genech (v rRNA, tRNA genech, př. 23S rRNA u Salmonellatyphimurium) • Nukleoid neoddělen od cytoplazmy membránou – translace přímo navazuje na transkripci • 1 replikační počátek/genom • Haploidní genom • Eukaryota: • Větší genomy, nižší hustota genů (klastry genů a genové pouště) • Velké procento genu tvoří introny (kombinace exonů, rekombinace, snížení rizika mutací) • Rozsáhlé intergenové oblasti (unikátní/repetice), větší počet regulačních sekvencí • Jaderná membrána, posttranskripční úpravy pre-mRNA (hnRNA) a teprve poté přesun do cytosolu • Více replikačních počátků/genom • Diploidní/polyploidní genom
Paradox C hodnoty C-hodnota = obsah DNA v haploidním genomu (bp, pg) Paradox C-hodnoty = neexistuje jednoduchý přímý vztah mezi velikostí genomu a biologickou (genetickou) komplexitou organizmu Totéž platí i pro G-hodnotu (počet genů) Nejmenší genom: Mycoplasma genitalium 500kb Největší genomy: např. mloci, nebo liliovité rostliny (velikost zhruba 100x lidský genom)
Osekvenované genomy Další příklady osekvenovaných genomů: Prokaryota: E.coli Viry: SARS Rostliny: rýže (Oryza sativa, 2002), huseníček rolní (Arabidopsis thaliana, 2000), kukuřice setá (Zea mays), topol chlupatoplodý (Populus trichocarpa, 2006), mech Physcomitrella patens (2008), vinná réva (Vitis vinifera, 2007) , papája (Carica papaya, 2008), čirok (Sorghum bicolor, 2009) Hmyz: D. melanogaster (2000), komár Anopheles, včela medonosná (2004), bourec morušový (91% v 2004) Ryby: zebřička (Danio rerio), čtverzubec fugu (Takifugu rubripes, 2002), čtverzubec černozelený (Tetraodon nigroviridis, 2004) Obojživelnící: drápatka Xenopus tropicalis Ptáci: kur bankivský (Gallus gallus, 2004) Savci: myš (2002), prase (2005), šimpanz (2005), pes (boxer)
Sekvenování = Stanovení sekvence nukleotidů v molekulách NK Dideoxy – metoda terminace řetězce (Sanger a Barell, 1977) Metoda chemického štěpení (Maxam a Gilbert, 1977) Next generation sequencing: 454 pyrosekvenování, SOLiD, Solexa, Helicos, Ion-Torrent… Sekvenování dvou lidských genomů (automatické sekvenátory) celková cena 300 milionů dolarů, každý chromozom se sekvenoval několik týdnů x pyrosekvenovaný genom J. Watsona byl celý hotový za 2-3 týdny, celková cena 100 tisíc dolarů
Sekvenování Dideoxy – metoda terminace řetězce (Sanger a Barell, 1977) + -
Sekvenování Dideoxy – metoda terminace řetězce - současnost
Lidský genom • Lidský jaderný genom (projekt HUGO = Human Genome Organization, založen 1990) • Publikace 2001, zpřesnění 2004, 2006, shotgun metoda • 25,2 % geny (1,2% exony, 24% introny); • 22 287 genů kódujících proteiny • (odhad z roku 2004; dnes okolo 21 000 genů) • 21-22% mezigenová DNA (heterochromatin, regulační sekvence, pseudogeny, genové fragmenty) • 50% genomu jsou repetitivní sekvence (tandemové x disperzní; vysoce x středně repetitivní) • Tandemové: centromery – satelitní DNA, telomery, mikrosatelity, minisatelity, rDNA klastry…) • Disperzní: většina transponovatelných elementů - 45% genomu!!!!! Celera Genomics (Venter), 2001 Mezinárodní Konsorcium, 2001
Lidský genom Lidský mitochondriální genom 16,6 kbp, 37 genů 22 genů pro rRNA, 2 pro tRNA, 13 pro proteiny dýchacího řetězce (ty jsou syntetizovány na mitochondriálních ribozomech, zbytek je kódován v jádře, syntetizován na ribozomech v cytosolu a pak exportován do mitochondrií) - Původ mitochnondrií z α – proteobakterií (Ricketsia) - endosymbióza
Lidský genom Dnes podle ENSEMBL: http://www.ensembl.org/Homo_sapiens/Info/StatsTable?db=core • Databáze sekvencí DNA • GenBank (USA)– dnes provozována Národním centrem pro biotechnologické informace (NCBI = National Center for Biotechnology Information), která je součástí Národní lékařské knihovny (NLM = NationalLibraryofMedicine); www.ncbi.nih.gov • EMBL (EuropeanMolecular Biology Laboratory) • DDBJ (DNA DataBankof Japan) • Dnes propojeny dohromady všechny; v databázích se dají hledat sekvence podobné zadaným sekvencím (BLAST = Basic LocalAlignmentSearchTool) • http://www.ncbi.nlm.nih.gov/Genomes/
Genomika = Studium celých genomů (včetně vývoje metod potřebných pro toto studium) • Oblasti genomiky • 1) strukturní: studium struktury genomu; konstrukce genomových sekvencí, identifikace a lokalizace genů, tvorba map (sekvenování…) • 2) srovnávací: studium evoluce genomu; srovnávání genových sekvencí mezi různými genomy (shody a rozdíly, syntenní mapy) → stanovení evolučních vztahů mezi organismy, studium evoluce chromozómů, identifikace genů, regulačních sekvencí, určení funkce genových produktů, lokalizace genů • 3) funkční: studium biologické funkce genů, jejich produktů, a jejich regulace; analýza transkriptomu a proteomu; z toho pak vznik transkriptomiky (microarrays, chromatinová immunoprecipitace, real-time PCR, PCR s reverzní transkripcí, RACE, SAGE…) a proteomiky (hmotnostní spektrometrie, krystalografie, dvouhybridová analýza, lokalizace proteinů pomocí GFP apod..)
Třídění sekvencí v genomu Unikátní (jedinečné) sekvence: 1-10 kopií/haploidní genom (geny, regulační oblasti, pseudogeny) Repetitivní sekvence: >10 kopií/ haploidní genom rozlišení na základě kinetiky DNA renaturace v 60. let. 20. st. – R. Britten, D. Kohne
Repetitivní sekvence Vysoce repetitivní sekvence – více než 100 tisíc kopií / n Satelitní DNA (samostatný pruh při hustotní centrifugaci v gradientu CsCl), obvykle se nacházejí v tandemu (za sebou)
Repetitivní sekvence Vysoce repetitivní sekvence – Satelitní DNA α - Satelitní DNA– shluky na 1/několika málo místech genomu (alfa-satelit v lidských centromerách – 171 nt motiv) Minisatelitní DNA = VNTR (variable number of tandem repeats), 10-100bp motiv, několik desítek – stovek kopií za sebou; hodně v subtelomerických oblastech; patří sem i telomery MikrosatelitníDNA= STR (short tandem repeats), 2-10bp motiv, několik tisíc kopií v tandemu za sebou, nejčastěji dinukleotidové (př. CACACACACACACA…) – ještě více polymorfní než VNTR (slippage DNA polymerázy nebo nerovnoměrný crossing over – vznik nových alel); vyjímečně i v genech – př. (CAG)n u Huntingtonovy chorey, expanze tri-nukleotid. repetic Využití repetitivních sekvencí ve forenzní genetice a v molekulární taxonomii (mikrosatelity, RAPD, RFLP). * Mikrosatelity a minisatelity někdy brány jako středně repetitivní
Forenzní genetika - Usvědčení pachatele trestného činu - Identifikace mrtvých osob - Příbuzenské vztahy (paternita) - Paleogenetika • STR profil • 16 STR lokusů(nekódující oblasti DNA) • Polymorfní • Každý člověk má individuální sestavu různých alel STR • Již 10 lokusů by stačilo na odhalení 1 osoby z populace 10 miliard lidí
STR – vznik nových alel 1) Nerovnoměrný crossing - over 2) Sklouznutí (slippage) DNA polymerázy během replikace Slippage: může být dopředu/dozadu; vytvoří se vlásenka - vyboulí se staré/nové vlákno DNA; nejdříve dojde k disociaci nového řetězce od templátu, při zpětné reasociaci nastává problém…
Repetitivní sekvence • Středně repetitivní sekvence: 10 - 100 tisíc kopií / n • Obvykle disperzní (rozptýlené) repetice • většina transponovatelných elementů (TEs) - disperzní • Multikopiové genové rodiny = geny pro ribozomální proteiny, aktin a myosin, histony, rRNA geny – jsou potřebné ve velkém množství – tandemové uspořádání Erythrinus erythrinus (Bertolo et al. 2010)
Transponovatelné elementy TEs = úseky DNA, které se mohou přemisťovat (přeskakovat) z jedné pozice hostitelského genomu do jiné (transpozice) Dle způsobu transpozice: Class I = Retrotranspozony Class II = DNA transpozony
Unikátní sekvence Geny, regulační sekvence, pseudogeny, nefunkční mutované geny, fragmenty, endogenní retroviry, počátky replikace … GEN = organizovaný úsek nukleové kyseliny projevující se a přenášející se jako základní jednotka dědičné (genetické) informace. Strukturní gen - kóduje polypeptid Gen kódující funkční RNA - kóduje tRNA, rRNA a řadu dalších RNA Gen jako regulační nebo strukturní oblast - promotory, enhancery, centromery, telomery ... Gen jako dědičná variabilní oblast nukleové kyseliny - satelitní polymorfní DNA (alely STR)... Přesná definice genu neexistuje…
Vývoj pojetí genu 1) J.G. Mendel (1866): gen je „jednotkový faktor“, který řídí specifický fenotypový znak jako např. barva květů hrachu (znovuobjevení roku 1900) Mendelovy zákony: 1) Zákon o uniformitě hybridů F1 generace po křížení homozygotních rodičů a identitě reciprokých křížení Křížíme-li dominantního homozygota s homozygotem recesívním, jsou jejich potomci F1 generace v sledovaném znaku všichni stejní. Nezáleží, zda je vloha zděděna od otce či od matky. 2) Zákon o čistotě a náhodné segregaci alel do gamet U heterozygota se dvě alely v průběhu tvorby gamet od sebe oddělují, segregují se (meióza); nestejnorodost F2 generace s fenotypovým štěpným poměrem 3:1; znovuobjevení recesivního fenotypu 3) Zákon o volné kombinovatelnosti alel Vlohy pro jednotlivé znaky jsou na sobě nezávislé
Vývoj pojetí genu Co Mendel nevěděl: neúplná dominance mnohotný alelismus intergenové interakce letalita (neživotaschopnost některých zygot) znaky vázané na pohlavní chromozomy mimojaderná dědičnost (mitochondrie – matroklinita); (u pohlavních chromozomů a mimojaderné dědičnosti neplatí identita reciprokých křížení) genová vazba (neplatí zákon o volné kombinovatelnosti alel) dědičnost kvantitativních znaků meiotický tah a genová konverze (porušují zákon o segregaci alel) epigenetická dědičnost, genomický imprinting horizontální přenos (alela není ani u jednoho z rodičů)
Vývoj pojetí genu • 2) A. Garrod (1909):1 mutantní gen = 1 metabolický blok • monogenní choroby u člověka (alkaptonurie, albinismus) • některé lidské choroby jsou způsobené „vrozenými chybami metabolismu“ jako výsledek ztráty určitého enzymu • 3) Thomas Hunt Morgan (1915) • - Polytenní chromozomy Drosophillamelanogaster(2n=8) • - geny jsou striktně lineárně uspořádány za sebou • geny jsou lokalizovány na chromozomech • geny, které jsou spolu na jednom chromozomu, nesegregují nezávisle na sobě (genová vazba)
Vývoj pojetí genu • 4) G.L. Beadle, E.L. Tatum (1940): 1 gen = 1 enzym • Auxotrofní mutanty plísně Neurosporacrassa (RTG nebo UV) • každá enzymově katalyzovaná chemická reakce je v organismu řízena jedním genem 5) L. Pauling, J.V. Neel, J.A. Beet (1949): 1 gen = 1 protein - studium srpkovité anémie, hemoglobiny se liší svými chemickými vlastnostmi (HbA, HbS); geny tedy nedeterminují jen enzymy 6) V. M. Ingram (1954-1957) 1 gen – 1 polypeptid - prokázal, že změna HbS je v globinu a ne v hemu - Proteinové komplexy z více podjednotek; enzymy složené z více polypeptidů kódovaných různými geny
Další vývoj představ o koncepci genu: 7) geny někdy kódují pouze RNA (tRNA, rRNA); 1 gen = 1 transkript 8) eukaryotické geny jsou přerušované introny + mají regulační sekvence (enhancery, silencery) 9) alternativní sestřih jeden gen může kódovat více polypeptidů (proteinové izoformy), jiná kombinace exonů/intronů nebo jejich částí
Vývoj pojetí genu 10) Genové segmenty u V(D)J rekombinace protilátek – variabilita protilátek, stavebnice segmentů 11) Geny uvnitř genů, překrývající se geny(viry) - genom fága ΦX174 (ssDNA, 5386nt; Φ = fí) kóduje 11 proteinů, které se ale nevejdou svou informací do celkové DNA viru - součet AMK z proteinů fága je 2300, ale dle genetického kódu 5386/3 má fág kapacitu jen na 1795 AMK! – jak to může kódovat? - Po sekvenování genomu odhaleno: Překrývající se geny - odlišné čtecí rámce téže molekuly DNA kódují různé proteiny, výjimka z nepřekryvnosti genetického kódu (možnost programovaného frameshiftu)
Uspořádání genů v genomu Prokaryota • Operony – geny jedné biosyntetické dráhy regulovány z jedné cis-regulační oblasti (operátoru) sousedící s promotorem; geny se přepisují dohromady (polycistronní transkript); pořadí genů = posloupnost biosyntetické dráhy Tryptofanový (Trp) operon Jacob a Monod – operonová teorie (1961) – laktózový (Lac) operon
Uspořádání genů v genomu Eukaryota • Každý gen má svoji vlastní regulační oblast – více možností regulace; monocistronní transkripty (ale C. elegans – 25% genomu dicistronní) Uspořádání genů možná není zcela náhodné- shluky genů s podobnými expresními profily Promotor: sekvence DNA, která určuje polohu transkripčního startu a směr transkripce, před genem; místo nasednutí RNA polymerázy Terminace transkripce: polyadenylační signál (u mRNA)
HOX geny • • specifický případ uspořádaného klastru genů eukaryot • Transkripční faktory, které řídí základní rozvržení těla během vývoje živočišných embryí podél ocaso-hlavní osy • regulovaný vznik struktur těla ve správném čase na správném místě • Každý obsahuje homeobox (homeodoména – vazba na DNA) • v roce 1923 Bridges a Morgan popsali mutaci bithorax u drozofily Normální moucha Mutace v genu bithorax– nadbytečný pár křídel místo končetin zvaných haltery (kyvadélka) Mutace v genu Antennapedia – nohy místo tykadel; nesprávná exprese genu Antennapedia v hlavičce
HOX geny Prostorové uspořádání HOX genů odpovídá tomu, v jakém pořadí budou exprimovány. Většina z nich je exprimovaná vysoce jen v některém stádiu (tmavší barva), později už tolik ne, jindy vůbec. U obratlovců jsou 4 HOX komplexy, každý na jiném chromozomu, ale když se přenesou do mouchy, tak částečně nahradí její HOX komplex. 4 HOX komplexy zřejmě vznikly duplikací hmyzích dvou klastrů.
Typy genů • Esenciální geny – nutné pro život (u člověka 5% genů, u E.coli 50%) • Housekeeping geny (udržovací geny) • - patří mezi esenciální geny, exprimují se v každé buňce bez rozdílu • - Kódují proteiny pro běžný provoz buňky • - geny pro tRNA, rRNA, ribozomální proteiny, histony, polymerázy, některé enzymy aj. • Tkáňově-specifické geny • Exprese jen ve specifických tkáních, v jiných buňkách umlčeny Průměrně velký lidský gen 27000 bp, ale pouze 1300 bp skutečně kóduje průměrně velký protein (430 aminokyselin u člověka)
Rekordy • Nejmenší geny • Microcin C7 • bakteriální antibiotikum microcin C7 • oligopeptid sestávající ze 7 AK • nesen plazmidem E. coli, má 27 nt a je přepisován spolu s dalším microcinovým genem, má samostatné vazebné místo na ribozóm umístěné před kodonem pro fMet • Největší gen • Dystrofin • 2.5 Mb dlouhý (0.1% genomu), 14kb mRNA • 79 exonů • 78 intronů (99,4% délky genu) • sestřihnutá mRNA = 14 000 nt • 8 promotorů • exprese ve svalech a mozku • poloha Xp21 • delece: Duchenneova muskulární dystrofie (1:3500 u mužů) nebo Beckerova MD (slabší příznaky, př. delece bez posunu čtecího rámce)
Lidský genom Odhady počtu protein kódujících genů u člověka 1997: ~100 000 2000: ~ 60 000 2001: 30 000 - 40 000 2004: 20 000 - 25 000, tj. ~ 1,2% genomu (když bereme v úvahu jenom exony) Resume: původní odhady zkresleny, nepočítalo se s alternativním sestřihem, alternativními promotory, alternativními poly-A signály atd.. Počty protein kódujících genů u jiných organismů myš 23 000 Drosophila 14 000 C. elegans 20 000 Arabidopsis 25 000 Resume: Člověk zhruba stejný počet genů jako C. elegans, ale vyšší počet alternativních transkriptů a post-translačních modifikací - vzniká až 5 x více různých proteinů u člověka
Lidský genom Šokující zjištění!! Zatímco, protein kódující geny tvoří u člověka jen asi 1,2% jaderného genomu, transkribováno je až 90% lidského genomu! - rRNA, tRNA, snRNA, snoRNA, miRNA, siRNA, piRNA - Alternativní transkripty - Antisense transkripty (>50% kódujících genů má antisense transkript) • Dlouhé nekódující RNA (např. Xist). • Transkribované pseudogeny
Pseudogeny Pseudogeny = nefunkční kopie genů. Sekvenční homologie se známými geny (ale rychle mutují). Mohou být transkribovány, mohou mít regulační fci? (antisense orientace – siRNA) Neupravené pseudogeny: vznik duplikacemi a následnými mutacemi genů, nebo reverzní transkripcí pre-mRNA a inzercí DNA kopie pre-mRNA do genomu (reverzní transkriptáza retrotranspozonů nebo retrovirů), také jako činnost Helitronů Upravené pseudogeny: vznik reverzní transkripcí mRNA bez intronů +inzerce cDNA do genomu, chybí regulační sekvence v DNA Vyřazené pseudogeny: vznik mutacemi v genech; předčasné stop kodony v mRNA apod. Pro započetí reverzní transkripce stačí poly-A –konec a ten má téměř jakákoliv mRNA
Evoluce genů 1) Evoluce genovou duplikací (Ohno, 1970) Duplikace části genu, celého genu (genová rodina), klastru genů, celého genomu (polyploidizace) - Po duplikaci genu si 1 kopie zachová původní funkci, druhá získá novou funkci (neofununkcionalizace) nebo (většinou) se stane pseudogenem 2) Exon shuffling– kombinace exonů, chimérické geny, činnost transponovatelných elementů (př. Helitrony 3) Horizontální přenos - př. u bakterií přenos genů rezistence na ATB pomocí plazmidů), přenos genů z endosymbiotické bakterie Wolbachia na hostitelské druhy hmyzu; gen syncitin u člověka (z genu env endogenního retroviru; úloha při formaci placenty); využití Ti plazmidu (tumor-inducing) zA. tumefaciens – schopnost včlenění do genomu rostliny (T-DNA) - transgenoze, Bt-kukuřice (protein z Bacillus thuringiensis – odolnost vůči škůdci) 4) Vnitrobuněčný přenos – z plastidů, mitochondrií
Homology genů Homolog (z řec. homos = stejný) - dá ještě dále třídit na: ortholog - homologické geny u různých druhů mající společného předka a většinou plní podobné funkce - odraz evoluce; př. hemoglobin člověka a krávy paralog - homolog odvozený genovou duplikací a následnou diverzifikací v rámci jednoho druhu; často mají odlišnou funkci – př. myoglobin, -hemoglobin, -hemoglobin u člověka xenolog - homolog získaný horizontálním přenosem - řada genů pro rezistenci k antibiotikům, ATPasy vakuolárního / archealniho typu u Gram+ bakterií, archealní lysyl-tRNA synthetasa u Borrellia burgdorferi) synolog - homolog v jednom organismu získaný po fúzi dvou nepříbuzných organismů - mitochondrie, chloroplasty v eukaryontní buňce) Genové rodiny = skupiny genů, které se během evoluce vyvinuly z jednoho společného předka a zachovaly si vysoký stupeň sekvenční podobnosti (stejné/různé funkce); pouze 76/2 200 genových rodin společných pro všechny organismy; člověk má 15 000 genů v genových rodinách
Příklad konvergence • Vznik genu pro nemrznoucí glykoprotein AFGP (antifreezeglycoprotein) u ryb žijících v polárních oblastech (odlišná forma v Antarktidě a Arktidě). • AFGP gen vznikl před 2,5 miliony let • přestože obě formy genů vznikly z různých genů, oba obsahují dlouhé úseky kódující tripeptidThr-Ala-Ala • Antarktický gen vznikl z genu pro trypsinogen amplifikací tripeptidové sekvence • Vznik arktického AFGP nemá nic společného s genem pro trypsinogen • Konvergentní evoluce – vznik u nepříbuzných organismů nezávisle na sobě, evoluce je dovedla v podobných prostředích k podobnému výsledku.
Zvětšování genomů 1) Duplikace - polyploidizace (duplikace celého genomu); zejména u rostlin (ale u savců př. osmáci jsou oktaploidní) - segmentální duplikace (duplikace chromozomových segmentů) - tandemové duplikace (duplikované geny / segmenty řazeny za sebou na stejném chromozómu) - genové (ektopické) duplikace (duplikované geny / menší skupiny genů nejsou řazeny za sebou) 2) Akumulace transponovatelných elementů – 80% genomu u kukuřice, 50% genomu rýže… 3) Inzerce cizorodých sekvencí
Zmenšování genomů Př. homologická rekombinace mezi různými kopiemi retroelementů v rámci jednoho řetězce DNA → musí to být rekombinace mezi přímými repeticemi!!, vyštěpí se kus chromozomu a je ztracen (nemá centromeru) markerem po deleci jsou solo LTR sekvence
Děkuji za pozornost.. Prezentace částečně vychází z přednášek: Genetika (Holá, Kočová), Evoluční genetika (Munclinger, Reifová), Forenzní genetika (Vaněk, Šimková), Základy molekulární biologie (Pospíšek), Virologie (Forstová), Genetika rostlin (Holá, Kočová, Rothová)