Download

1 / 14
140 likes | 321 Views
Cholinergic induction of network oscillations at 40Hz in the hippocampus in vitro. Fisahn et. al, Nature Vol 394 (1998), pp. 186. Un modelo.

E N D
Cholinergic induction of network oscillations at 40Hz in the hippocampus in vitro Fisahn et. al, Nature Vol 394 (1998), pp. 186 Un modelo En el siguiente trabajo modelamos las oscilaciones autosostenidas a 40 Hz.medidas in vitro a través de la técnica de registro de campo en una red de células piramidales del hipocampo de rata. El modelo consta de un sistema de ecuaciones que modela la dinámica de la población de células piramidales y gabaergicas, acoplado a un modelo de neurona basado en Hodgkin-Huxley con un término asociado a la contribución de las células vecinas. Ajustando los parámetros del modelo se observaron las oscilaciones y se reprodujeron los experimentos farmacológicos. Julieta Sztarker - Hernán E. Grecco Física de Sistemas Biológicos Julio de 2003
Introducción: En el trabajo “Cholinergic induction...” se observa la aparición de oscilaciones de 40 Hz en la red de células piramidales del hipocampo de rata ante la estimulación con carbacol (agonista de los receptores de acetilcolina) in vitro. La aparición de este tipo de oscilaciones sincrónicas a 40 Hz se observa también registrando la actividad de poblaciones de neuronas in vivo. El estudio del hipocampo es particularmente interesante ya que se sabe juega un rol central en tareas relacionadas con la memoria. En este trabajo se emplean registros de campo local (con un electrodo extracelular ubicado cerca del tejido) en rodajas de hipocampo ubicadas en un baño: V
Dicho baño permite la aplicación transitoria de diversos fármacos para ver cual es el efecto de los mismos sobre la actividad poblacional de las neuronas. En particular observaron que ante la aplicación de carbacol se producían oscilaciones autosostenidas de 40 Hz de frecuencia: El panel inferior muestra el espectro de Fourier para el trazo control (línea inferior) y para la aplicación de carbacol (trazo superior) en el que se puede observar el pico de frecuencia predominante a 40 Hz. Utilizando bloqueantes específicos para los distintos subtipos de receptores se observó que son los receptores muscarínicos M1 los involucrados en este fenómeno.
Los receptores de GABA están acoplados a un canal de Cl-, que se abre al pegar una molécula de GABA hiperpolarizando a la célula. Se dice entonces que el GABA es un neurotransmisor inhibitorio ya que aleja a la célula del umbral de disparo de un potencial de acción. Al colocar en el baño un antagonista de los receptores de GABA, la bicuculina, se ve que se eliminan las oscilaciones: Este resultado demuestra que sinapsis inhibitorias están involucradas en la sustentación de las oscilaciones. Para confirmar el rol de las sinapsis inhibitorias en este proceso, utilizaron el fármaco, pentobarbitona, el cual cambia la cinética del IPSP (potencial inhibitorio postsináptico), enlenteciendo la fase de subida: La frecuencia de oscilación disminuye alagregar pentobarbitona, indicando que son las sinapsis inhibitorias las que regulan la frecuencia de la oscilación.
El glutamato es un neurotransmisor excitatorio como la acetilcolina, que al interactuar con sus receptores provoca la apertura de un canal permeable al Ca2+, que alentrar en la célula la depolariza, es decir la lleva más cerca del umbral de disparo de un potencial de acción. Para saber si el sistema glutamatérgico estaba involucrado en la sustentación de las oscilaciones aplicaron al baño bloqueantes específicos para los distintos subtipos de receptores. Encontraron que el fármaco NBQX, antagonista de los receptores de glutamato no NMDA eliminaba las oscilaciones generadas por aplicación de carbacol. Este resultado demuestra que sinapsis excitatorias glutamatérgicas están involucradas en la sustentación de la oscilación. Los resultados indican que sinapsis de tipo glutamatérgicas y gabaérgicas participan en la sustentación de las oscilaciones de 40 Hz. Sin embargo, al haber hecho estos estudios utilizando registros de campo, no podemos saber si los dos tipos de sinapsis coexisten en la misma neurona o si se encuentran en neuronas distintas.
Para dilucidar este punto realizaron registros intracelulares de tipo patch clamp en las células piramidales. Utilizaron la configuración de voltage clamp, en la cual se inyecta corriente en la célula para compensar las corrientes producidas y así mantener fijo el voltaje de membrana de la célula al voltaje deseado. Lo que se mide es dicha corriente en función del tiempo. Al fijar el voltaje de membrana a -70 mV, sólo observamos las corrientes depolarizantes (EPSC: corriente excitatoria postsináptica) ya que los iones hiperpolarizantes (K+ y Cl-) se encuentran muy cerca de su potencial de equilibrio. Al fijarloa +20 mV, sólo vemos las corrientes hiperpolarizantes (IPSC: corriente inhibitoria postsináptica ) ya que los iones depolarizantes (Na+ y Ca2+) se encuentran muy cerca de su potencial de equilibrio. Este resultado nos indica que la misma neurona piramidal recibe los dos inputs sinápticos.
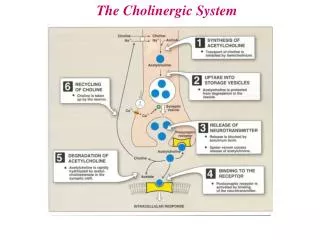
Interneurona gabaérgica Célula piramidal Considerando todos los resultados anteriores, los autores proponen el siguiente modelo de cómo serían las conexiones entre las neuronas en el hipocampo:
Modelo de célula piramidal Cada célula puede ser disparada por la presencia de Carbacol con baja probabilidad. Los pulsos de Carbacol fueron introducidos en el modelo de neurona de Hodgkin-Huxley (HH) como pulsos de corriente. Para simular la aleatoriedad, se generó un tren de N pulsos de igual alto cuyos centros eran generados al azar dentro de la duración del experimento y con anchos distribuidos uniformemente alrededor de un valor medio. Si bien cada pulso entregado es subumbral el factor azar hace posible que varios de estos pulsos coincidan en un tiempo reducido (simulando el fenómeno de sumación temporal) y hagan a la neurona disparar. Sin embargo la actividad de una neurona depende también de la actividad de las neuronas vecinas. Para incorporar este parámetro incluimos un termino aditivo al potencial de membrana dentro del modelo HH proporcional al voltaje medio de la población: donde V es el voltaje medio del campo y v es el voltaje de membrana de la celula relativo al voltaje de reposo.
k1 Modelo de la actividad media k2 k3 Teniendo en cuenta que la mayoría de los experimentos realizados en el trabajo de Fizhan et al son registros de campo en los que no se mide el potencial de membrana de cada célula sino un voltaje medio que refleja el estado del sistema, no tiene sentido hacer un modelo detallista de la red considerando todas las interconexiones entre las neuronas, sino describir el comportamiento macroscópico del sistema. Siguiendo el modelo de Fizhan et. al., las células piramidales excitan a otras células excitatorias y a las gabaergicas. Estas últimas inhiben a las piramidales. Donde V es el voltaje medio de la población de células piramidales e I es el voltaje medio de la población de células gabaergicas. Dado que nos interesa estudiar fenómenos colectivos, colapsamos cada sinapsis en un único proceso caracterizado por las constantes K1 y K2. El ultimo termino de la ecuación para V representa la influencia de cada célula individual. Para incluir el aporte de varias células individuales al modelo, sumamos 15 corridas independientes del modelo para cada experimento. Estas corridas se inician con un pulso a t = 0 para sincronizar las oscilaciones.
Resultados Ajustando los valores de los parámetros, pueden reproducirse los resultados experimentales en cuanto a la sustentación del proceso y su frecuencia. A la izquierda se observa el resultado del modelo y su espectro de Fourier acordes con el experimento (derecha).
La bicuculina inhibe los receptores GABA k1 k2 k3 k2=0 Variando el valor de los parámetros K1 y K2 pudimos reproducir los resultados farmacológicos. Por ejemplo el aplicar bicuculina (antagonista gabaergico) al medio es comparable a desconectar las neuronas gabaergicas de las piramidales, es decir hacer K2=0. Al igual que el registro de campo la salida del modelo no presenta oscilaciones a ninguna frecuencia típica, como se observa en el espectro de Fourier.
k1 k2 k3 El efecto de la pentobarbitona se puede simular reduciendo el valor de K2, por ejemplo a la mitad. La pentobarbitona hace mas lento el IPSP Disminuir k2 El corrimiento en frecuencia se puede ver en el espectro de Fourier, Grafico de la izquierda negro = (k2 original), rojo= (k2 original)/2. Grafico de la derecha trazo gris = original, negro = luego de la aplicación de pentobarbitona.
k1 k2 k3 La aplicación de NBQX (antagonista glutamatergico no NMDA) en el medio, se puede modelar haciendo K1=0. NBQX inhibe receptores de glutamato k1 =0 Nuevamente el modelo refleja la ausencia de oscilaciones sincronizadas cuando el NBQX esta presente.
Conclusiones Nuestro modelo presenta resultados acordes a los medidos por Fizhan et. al. Variando los parámetros k1 y k2 se pueden modelar los procesos inducidos por los fármacos utilizados en el trabajo (supresión de las oscilaciones, cambio de frecuencia). En este caso, un modelo sencillo como el planteado, con pocos parámetros, variables macroscópicas y procesos simplificados, mostró ser suficiente para describir los comportamientos mas destacados de la red. De ser necesario el modelo podría complejizarse para explicar los resultados de otros experimentos realizados en la preparación. Considerando la dinámica de los fármacos (bicuculina, pentobarbitona, NBQX) sobre los respectivos receptores, se podría modelar k1 y k2 dependiendo de la concentración de los mismos. En la salida de nuestro modelo observamos que existe información importante (tiempos típicos, etc) en los transitorios del sistema, que no están presentes en el trabajo (solo muestran los estados estacionarios) y que seria interesante de comparar.