Download

1 / 16
160 likes | 302 Views
Modular proteins I. Level 3 Molecular Evolution and Bioinformatics Jim Provan. Patthy Sections 8.1.1 – 8.1.3. Protein domains. Folded structures of proteins that are larger than 200-300 residues generally consist of multiple structural domains :

E N D
Modular proteins I Level 3 Molecular Evolution and Bioinformatics Jim Provan Patthy Sections 8.1.1 – 8.1.3
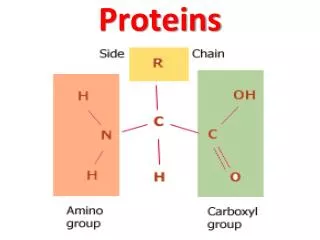
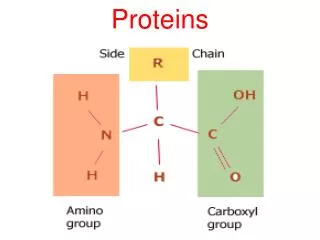
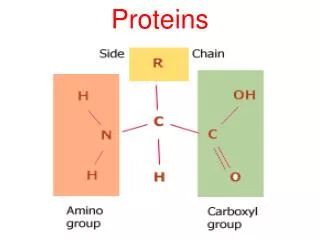
Protein domains • Folded structures of proteins that are larger than 200-300 residues generally consist of multiple structural domains: • Compact, stable units with a unique three-dimensional structure • Interactions within a domain are more significant than those between domains • Fold independently i.e. structural domains are also folding domains • If domain performs distinct function which remains intact in the isolated domain, then it is also a functional domain • Many multidomain proteins are homomultimeric i.e. contain multiple copies of a single type of structural domain: • Arisen through internal duplication of complete domains • Fate of domains determined by similar rules to paralogous genes
Protein domains • Many multidomain proteins are heteromeric: • Example is plasminogen activator where a trypsin-like serine protease is joined to kringle, finger and EGF domains • May occur by fusion of two or more genes (chimeric proteins) • Also known as modular proteins, with domains known as modules • Certain modules occur in a wide variety of hetero- and homomultimeric proteins: • Suggests mechanisms to facilitate duplication and dispersal • “Building blocks” of different types of multidomain proteins are known as mobile protein modules • Frequency of transfer and incorporation into new protein reflects fixation probability
Modular assembly by intronic recombination • Discovery of introns provided potential new mechanisms for protein evolution: • Gilbert suggested that recombination within introns could assort exons independently • Idea of rapid construction of novel genes from parts of old ones led to the formulation of the exon-shuffling hypothesis • According to “introns early” theories, all extant genes were constructed from a limited number of exon types • Under the “introns late” theory, intronic recombination and exon shuffling could not have played a major role in the assembly of the earliest genes • Original theory was that exons corresponded directly to modules and/or structural motifs
Problems with the “introns early” hypothesis • In the case of many genes, no obvious correspondence was observed between protein structure and intron location • It is now known that introns can also be inserted into genes i.e. structure of a gene may not be its original structure • Introns suitable for exon shuffling did not originate until a relatively late stage of eukaryotic evolution • Exon shuffling has only been conclusively demonstrated in “young” proteins unique to higher eukaryotes • Only a special group of exons, the “symmetrical” modules, are really valuable for exon shuffling. Intron phase distribution is also a crucial factor.
Self-splicing introns • Group I introns: • Reaction requires only a guanine nucleotide cofactor: • Provides a free 3’-OH group that attacks the 5’ splice site • 3’-OH generated at the end of the upstream exon • Second transesterification joins the two exons • Crucially depends on folded structure of the intron itself • Group II introns: • Does not require an external cofactor: 2’-OH of an adenine within the intron cuts the 5’ splice site • 2’5’ phosphodiester bond (branch site) forms the lariat structure • Although folding is still crucial, chemistry, sequence of events and lariat formation are similar to nuclear spliceosomal introns
Spliceosomal introns • Spliceosomal introns are only spliced in the presence of a complex of specific proteins and RNA known as a spliceosome • Majority of intron is unimportant: as long as the 5’ and 3’ splice sites and the branch site are conserved, splicing can take place: • Large insertions into spliceosomal introns, or deletions do not affect splicing efficiency • Chimeric introns, containing the 5’ end of one intron and the 3’ end of another, are also properly spliced • Mutations (directed or otherwise) in these regions lead to aberrant splicing • Spliceosomal intron plays a minor role in its own splicing: the actual spliceosome complex is more important
Evolution of spliceosomal introns • Both group I and group II self-splicing mechanisms resemble spliceosome catalysed splicing: • Initial step is attack by ribose hydroxyl group on 5’ splice site • In each case, reactions are transesterifications where phosphate moieties are retained in products • In group II and spliceosomal introns, intron is released as a lariat • Accepted that spliceosomal-catalysed splicing evolved from group II self-splicing introns : • Key step was transfer of catalytic role from intron to other molecules • Formation of spliceosome gave spleceosomal introns structural freedom as they no longer had to fulfil the catalytic function • Generally found only in nuclear genomes of higher eukaryotes (plants, animals and fungi)
Intron loss • Plays a significant role in changing exon-intron structure of genes • Introns may be eliminated through mechanism that gives rise to processed genes (retroposition) • Reverse transcription can also lead to loss of only some introns: • Reverse transcription of perfectly spliced mRNA and recombination with the functional gene: mutates original gene • Partially processed pre-mRNA could give rise to a semi-processed gene: generates a new paralogue
Exon shuffling via recombination in introns • Believed that insertion of exons may occur by the same mechanism as insertion of introns: • Exon shuffling may be a consequence of the occasional inclusion of exon sequences in the insertion cycle of introns • Alternative splicing (exon skipping during splicing) may yield exons with flanking introns • If such a composite is inserted into the genome by the same mechanism that inserts single introns (reverse splicing) we have exon shuffling • Key difference between intronic recombination model and retrotransposition model: • In first case, insertion occurs into a pre-existing intron of same phase as introns flanking exon • Retrotransposition model does not have this requirement
S P G S G P K S G K P Evolution of urokinase
S G K P F S F G K P K module duplication S F G K K P Evolution of tissue plasminogen activator
S G K P Duplication of G module S G G K P F S G F G K P FN2 S FN2 G F G K P Evolution of Factor XII