Download

1 / 51
640 likes | 1.87k Views
Introduction to Enzymes Properties, active sites, specificity, rate acceleration Rates Free energy of activation, Arrhenius and Erying’s equations Kinetic concepts First order, second order reactions Steady state enzyme kinetics Km, kcat, kcat/Km Transient state kinetics

E N D
Introduction to Enzymes Properties, active sites, specificity, rate acceleration Rates Free energy of activation, Arrhenius and Erying’s equations Kinetic concepts First order, second order reactions Steady state enzyme kinetics Km, kcat, kcat/Km Transient state kinetics Rapid kinetic methods, kinetic simulations
Enzymes catalyze all the reactions of the living cell Nuclear Mitochondrial Ribosomal Microsomal Lysosomal Cytoplasmic
ENZYMES ARE PROTEINS James Sumner used crystallography (Urease) to establish that enzymes are proteins. Awarded Nobel prize in 1946. Polypeptide chain of amino acids Folded protein Not all proteins are enzymes
Protein enzymes have active sites Enzyme active sites are located in clefts Acetyl choline esterase Lysozyme
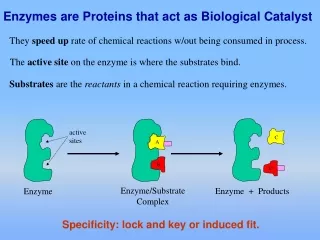
The Active site is a small part of the enzyme, a pocket, that is lined with a few catalytic amino acids that participate in binding and catalysis • Active sites are clefts or crevices that usually exclude water (unless it is a reactant). The nonpolar character of the active site enhances the reactivity of any polar residues that contribute to the active site • Substrates are bound to enzymes by many weak interactions; i.e. hydrogen bonds, electrostatic, Van der Waals and hydrophobic interactions Features of Enzyme Active Sites
Catalytic residues • Direct involvment in the catalytic mechanism. • Exerting an effect on another residue or water directly involved in the catalytic mechanism • Stabilization of a proposed transition state intermediate • Exerting an effect, electrostatic, on substrate or cofactor aiding catalysis
Active site cleft • Specific amino acids come together to form the active site. • substrates bind in the active site cleft through noncovalent interactions: • Electrostatic: ionic, dipole-dipole • H-bonds • Hydrophobic H-bonds ionic Substrate binding is reversible
The effect of DpH on enzyme activity • pH can have two effects on an enzyme: • At extremes of pH, the enzyme is irreversibly denatured and activity is permanently lost • pH can also reversibly alter the kinetics of an enzyme by affecting the ionization of enzyme functional groups involved in catalysis or substrate binding or by altering the substrate’s ionization state. Decline in activity results from an ionization event & NOT irreversible denaturation Stability Curve: activity at optimal pH after incubation at indicated pH value Decline in activity results from irreversible denaturation Activity Curve: direct assay in buffer at indicated pH
SPECIFICITY Narrow (absolute) or broad Chymotrypsin and glucokinase have absolute specificities Alcohol dehydrogenase has a broad specificity CH3CHO + NADH CH3CH2OH + NAD+ ethanol, methanol, propanol
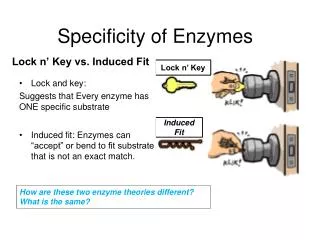
SPECIFICITY Models that explain enzyme specificity • Fischer’s lock and key model • “In order to be able to act chemically on one another, an enzyme and its substrate must fit together like a lock and key.” • Emil Fischer (1852-1919) • Awarded Nobel prize in chemistry, 1902
Fischer’s Lock and Key concept enzyme enzyme substrate Led medicinal chemists to design substrate-like inhibitors as potential drugs enzyme Why does the enzyme not catalyze reactions on smaller molecules?
Glucokinase ATP + glucose ADP + glucose-6-phosphate
Specificity galactose Glucose binding to glucokinase
SPECIFICITY Hexokinase or Glucokinase fast ATP + glucose ADP + glucose-6-phosphate In the cell, the concentration of H2O is much higher than glucose slow ATP + H2O ADP + phosphate
Specificity Induced fit theory Daniel Koshland "the substrate may cause an appreciable change in the three-dimensional relationship of the amino acids at the active site" “hand in a glove” rather than “lock in a key” Specific substrate binding induces a conformational change that strengthens binding and activates the enzyme
Binding of glucose induces a large conformational change The two lobes swing together by 12 A Glucose is completely engulfed by the protein active
Enzymes • General properties of enzymes • Proteins • Catalyst • Higher reaction rate • Milder reaction conditions • Specificity • Capacity for regulation
Reactions proceed spontaneously if there is a decrease in free energy, G. But, “spontaneous” does not imply that the reaction occurs at a measurable rate. C12H22O11 + 12 O2 12 CO2 + 11 H2O -5693 kJ/mol Who has seen a flash fire in a sugar bowl? In fact, most reactions are slow and require catalysts such as enzymes to proceed at reasonable rates. Reaction Rates
Non-enzymatic rates • How do the rates of enzyme-catalyzed reactions compare to uncatalyzed reactions? • Some enzyme-catalyzed reactions are so slow in the absence of an enzyme catalyst that their half-lives begin to approach the age of the earth itself!
Rate Enhancements (kcat/knon) Peptide hydrolysis Uncatalyzed: 10-10 s-1 Catalyzed: 103 s-1 Rate acceleration: = catalyzed/uncatalyzed = 1013 What is the basis for rate acceleration?
Mechanisms for Stabilizing Transition States and Accelerating Rates
Rate Acceleration by Proximity Bringing Two Substrates into Close Proximity at the Active Site of an Enzyme
General acid-base catalysis • Mechanism for bringing about catalysis • Functional groups of enzymes that can act as general acid or base include: • -COO- of asp and glu (pKa ~ 4.5) • imidazole of histidine (pKa~7) • -SH of cysteine (pKa~8-9) • protonated amino groups of lysine (pKa~10-11) • N-terminal amino group of peptide (pKa~8)
General acid-base catalysis The uncatalyzed hydrolysis of an acetal leads to a transition state in which positive and negative charges develop across the C—O bond. This charge separation is quite unfavorable and leads to a very slow rate of reaction.
General acid-base catalysis Thegeneral acid catalyzed hydrolysis of an acetal stabilizes the developing negative charge on the oxygen of the acetal by transferring a proton from the general acid catalyst. Partial proton transfer from a general acid catalyst lowers the free energy of activation by stabilizing the developing negative charge in the transition state.
General acid-base catalysis The uncatalyzed attack of water on an ester leads to a transition state in which positive charge develops on the attacking water molecule and negative charge develops on the carbonyl oxygen. These developing charges are unfavorable in the transition state.
General acid-base catalysis The general base catalyzed attack of water on an ester allows the developing positive charge on the water oxygen to be stabilized by the transfer of its proton to the general base catalyst. Partial proton transfer to a general base catalyst lowers the free energy of activation by stabilizing the developing positive charge in the transition state.
General acid-base catalysis • Acid-base catalysis is termed “general” to distinguish it from specific acid-base catalysis. In specific acid-base catalysis, the catalyst is the proton or hydroxide ion itself.
Example: Enolase general base catalyst general acid catalyst 2-phosphoglycerate phosphoenolpyruvate ene-diolate intermediate
Covalent catalysis - Nucleophilic catalysis Common nucleophlic groups on enzymes & example enzymes: Cysteine thiol Serine proteases Cholinesterases Esterases Serine hydroxyl Thiol proteases G3P dehydrogenase Lysine amino group PLP-dependent enzymes acetoacetate decarboxylase Histidine imidazole phosphoglycerate mutase succinyl-CoA synthetase
Chymotrypsin Tetrahedral intermediate covalent intermediate nucleophile
Common Characteristics • all contain a catalytic triad - serine, histidine, and aspartate • all share an oxyanion hole
nucleophile This arrangement of Asp, His, and Ser residues in chymotrypsin has become known as the charge-transfer relay system or catalytic triad
Acid-base and Covalent Catalysis General base General acid: donates proton General base: accepts proton Serine forms a covalent bond with the substrate
General acid General base Covalent intermediate
Enzyme binds the transition state of the reaction it catalyzes with far greater affinity than it binds either the substrates or the products. Michaelis Complex Tetrahedral Intermediate
Metal ion catalysis • Two classes: metalloenzymes (Fe2+, Fe3+, Cu2+, Zn2+, Mg2+, Mn2+, or Co3+) and metal-activated enzymes (Na+, K+, Mg2+, or Ca2+) • Participate in reactions: • By binding substrates • By mediating oxidation-reduction reactions • By electrostatically stabilizing or shielding negative charges
Metal ion catalysis • Can make a reaction center more susceptible to receiving electrons because it can stabilize a developing negative charge on a transition state. (electrophilic catalyst)
Metal ion catalysis • Can stabilize developing negative charge on a leaving group, making it a better leaving group. (electrophilic catalyst)
Metal ion catalysis • Can increase the rate of a hydrolysis reaction by forming a complex with water, thereby increasing water’s acidity.
Basis for increased reaction rate Preferential Transition State Binding “I think that enzymes are molecules that are complementary in structure to the activated complexes of the reactions that they catalyzed, that is, the molecular configuration (of the activated complexes) is intermediate between the reacting substances and the products of the reaction” Linus Pauling, 1948